FIGURE 57-1. Comparison of the effects of surgical thyroparathyroidectomy with parathyroidectomy by cautery or surgery in the rat.
(Data from Hirsch PF et al, Endocrinology 73:244–251, 1963.)
Synthesis, Secretion, and Cells of Origin
CT is produced by the C cells of the mammalian thyroid, with its secretion dependent on serum calcium levels.5 Although the dominant site of production of CT in mammals is the thyroid C cell, the distribution of these cells throughout the thyroid gland varies considerably among mammalian species, and there is evidence that in some animals CT-producing cells might be found in other parts of the neck, including the thymus. In fish and in most birds, CT is produced by the ultimobranchial glands. Whereas in mammalian development the ultimobranchial bodies fuse with the posterior lobes of the developing thyroid to become the C cells, in submammalian vertebrates these bodies remain separate, and the ultimobranchial glands constitute a separate endocrine system. The CTs of ultimobranchial origin are highly potent in their actions upon mammalian targets. The physiologic significance of CT in fish and birds remains uncertain, although it is interesting to note that CT has been reported to suppress osteoclastic activity in the scales of freshwater and seawater teleosts.6 Although there is little doubt that increased serum calcium is an important secretagogue for CT in normal and malignant C cells, the exact mechanisms by which calcium provokes exocytosis of CT have not been fully elucidated. The same extracellular calcium–sensing receptor that mediates decreased PTH secretion from parathyroid cells7 is also found in C cells and is likely to represent the primary molecular entity through which C cells detect changes in extracellular calcium and control CT release.
Agents that elevate C cell cyclic adenosine monophosphate (cAMP) may stimulate CT secretion, since cAMP analogues have this effect both in vivo and in vitro. Probably the most important CT secretagogues, apart from calcium, are the gastrointestinal hormones. In the pig, gastrin appears to be an effective physiologic secretagogue, suggesting that CT may have a physiologic role postprandially as a hormone that assists uptake of calcium after a calcium-rich meal by preventing the efflux of calcium from bone into blood.8 Although there is some evidence for this role in pigs and rats, studies in humans remain to be performed. Other gastrointestinal hormones, including glucagon, cholecystokinin, and secretin, are also capable of promoting CT secretion. The gastrin analogue, pentagastrin, has been used clinically as a provocative test for CT secretion in patients with medullary carcinoma of the thyroid. Other hormones that influence calcium homeostasis may also directly or indirectly influence CT secretion. 1,25-Dihydroxyvitamin D3 (1,25[OH]2D3) administration has been reported to increase plasma CT levels; this was suggested to occur via specific thyroid C cell receptors for 1,25(OH)2D3, which modify secretion of CT.9 Both CT and 1,25(OH)2D3 levels are raised in pregnancy and lactation, leading to the suggestion that CT may act to protect the skeleton in the face of increased calcium demand by the fetus.
Serum and thyroid concentrations of CT increase markedly with age in the rat, in association with substantial increases in thyroid content of CT mRNA.10 In normal rats subjected to acute calcium stimulation in vivo, thyroid CT mRNA is increased. On current evidence it seems that calcium can stimulate both synthesis and secretion of CT by thyroid C cells.
CT secretion has been studied extensively in patients with medullary carcinoma of the thyroid, who have elevated CT levels (see later). However, circulating CT levels in normal human subjects are very low and their measurement requires sensitive and specific assays. The level of CT in normal human blood appears to be less than 10 pg/mL (3 picomolar). Circulating levels of CT are increased in several pathologic states, such as CT-secreting tumors.11,12 In addition to the CT monomer (≈3500 Daltons), high molecular weight forms circulate, and elevated levels of these molecules can be useful diagnostically in certain situations, such as acute pancreatitis13,14 and infection/inflammatory conditions.15 In fact, it has been reported that ProCT is toxic and that immunoneutralization with immunoglobulin (Ig)G that is reactive to this molecule significantly improves survival in animal models of sepsis.16
Chemistry
The CT sequence has been determined for many species, the common features being that it is a 32 amino acid peptide with a carboxyterminal proline amide and a disulfide bridge between cysteine residues at positions 1 and 7.17 Based on their amino acid sequence homologies, the different CTs (Fig. 57-2) are classified into three groups: (1) artiodactyl, which includes porcine, bovine, and ovine; (2) primate/rodent, which includes human and rat CT; and (3) teleost/avian, which includes salmon, eel, goldfish, and chicken. The common structural features of the CT molecule contribute importantly to biological activity, with the standard assay that has been used since the discovery of CT being one that measures the hypocalcemic response in young rats. Subsequently, receptor-based assays have been used also, and structure/function relationships are largely shared in these various assays. The order of biological potency of the CTs is, in general, teleost≥artiodactyl≥human, although absolute biological activities vary considerably among CT receptors of different species and receptor isoforms within species. Studies of substituted, deleted, and otherwise modified CTs have provided considerable information regarding structure/activity relationships of the CT molecule, showing, for example, that the ring structure serves to stabilize the molecule. The disulfide bridge of the ring can be chemically substituted by an N-N bond, as in aminosuberic eel CT, and this modification yields an extremely stable and fully potent CT variant.18 The sequence differences among species are concentrated in the middle portion of the molecule, and these differences contribute to the wide variations in biological potencies. However, the outcomes of studies of structural requirements for biological activity have varied with the different biological assays used, and the type of receptor used is able to profoundly influence the results. For example, residues in the carboxyterminal half of salmon CT are more important for binding competition with the two rat receptor isoforms and the human receptor, whereas residues in the aminoterminus are more important for interaction with the porcine receptor.5

FIGURE 57-2. Alignment of CT sequences from different species. The shaded amino acids indicate identity with salmon CT. All CTs have a disulphide-bridged loop between cysteines (C) at position 1 and 7, a glycine at position 28, and a proline amide at position 32. Amino acids 4, 5, and 6 are also conserved across all species.
Calcitonin Gene
As with other hormonal peptides, CT is synthesized as a larger precursor molecule, which is processed by cleavage and amidation before secretion. CT is synthesized as a large molecular weight precursor (136 amino acids), with a leader sequence at the aminoterminus that is cleaved during transport of the molecule into the endoplasmic reticulum. A potentially important posttranslational modification of CT is that of glycosylation. It had been noted that the tripeptide sequence, Asn-Leu-Ser, found within the aminoterminal ring structure of CT is invariate among the CTs of different species. This sequence is an acceptor site for N-linked glycosylation. This, together with evidence for glycosylation of tumor CT, led to detailed studies showing that the CT precursor is indeed a glycoprotein, and that the only N-linked glycosylation site in the entire precursor was within the CT portion itself.19 The biological significance of CT glycosylation has yet to be determined.
The complete sequences of the cDNA for human, rat, mouse, chicken, sheep, dog,20 and various species of fish CTs and the DNA sequence of the full human CT gene have been determined.21–23 These show that the hormone is flanked in the precursor by N- and C-terminal peptides, but the biological significance of these peptides is unknown. The human CT gene has been located in the p14-qter region of chromosome 11.24
ALTERNATIVE GENE PRODUCT—CGRP
The CT gene transcript actually encodes a second distinct peptide known as CT gene-related peptide (CGRP), which is produced by tissue-specific alternative splicing of the gene (Fig. 57-3). The mature CGRP and CT mRNAs predict proteins that share sequence identity in the aminoterminal regions, but in the carboxyterminal regions the nucleotide sequences are almost entirely different. The mature, secreted 32 and 37 amino acid CT and CGRP peptides, respectively, result from cleavage of both aminoterminal and carboxyterminal flanking sequences at specific cleavage sites, as depicted in Fig. 57-3.24 CT mRNA is found largely in the thyroid, and CGRP mRNA is found primarily in the nervous system.25 However, aberrant expression of CGRP may be seen in medullary thyroid carcinoma.26 Two different CT/CGRP genes, α and β, have been identified in man and rat.
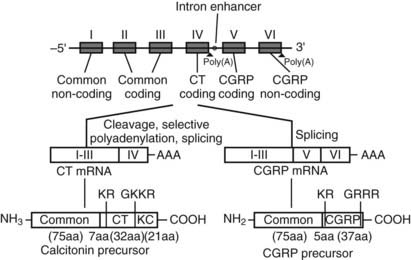
FIGURE 57-3. Organization of the CT/CGRP gene illustrating alternative patterns of processing of the primary transcript and subsequent protein processing. The exons are denoted I through VI in Roman numerals, introns are represented by a single line (not to scale).
Processing of the pre-mRNA to the CT mRNA transcript involves usage of exon 4 as a 3′-terminal exon with concomitant polyadenylation at the end of exon 4. Processing to produce the CGRP mRNA involves the exclusion of exon 4 and direct ligation of exon 3 to exon 5, with polyadenylation at the end of exon 6. The hCT/CGRP exon 4, like many differentially incorporated exons, has been characterized as having weak processing signals. Weak differential exons are frequently associated with special enhancer sequences that facilitate exon recognition in the presence of accessory factors that bind to the enhancer. Indeed, such an enhancer, located in the intron downstream of exon 4, has been described for the CT/CGRP gene. In addition, sequences within exon 4 are necessary for the inclusion of exon 4.27
Physiology—Bone and Kidney
The physiologic role of CT is not fully understood. It is currently viewed as an inhibitor of bone resorption, whose function is to prevent bone loss at times of stress on skeletal calcium conservation, particularly pregnancy, lactation, and growth. Earlier concepts of CT as a regulator of extracellular fluid calcium are probably relevant only in young and growing animals, in which rapid bone modeling and turnover are required for development of the skeleton. In the rat, for example, which is used for the in vivo assay of CT, the calcium-lowering effect of the hormone is less marked with increasing age of the animal (Fig. 57-4).28 However, the ability of CT to counteract the effects of a calcium load was shown not to be impaired in older animals, at least in the rat29—an observation that has not been explained and that has not been extended to other species.
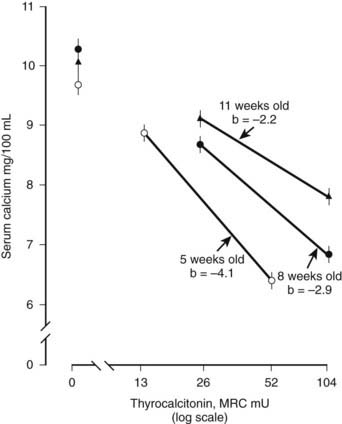
FIGURE 57-4. Decreasing hypocalcemic response to calcitonin with increasing age of the rat.
(Data from Cooper CW et al, Endocrinology 81:610–617, 1967.)
In normal adult human subjects, even quite large doses of CT have little effect on serum calcium levels. In those subjects in whom bone turnover is increased (e.g., in thyrotoxicosis, Paget’s disease), CT treatment acutely inhibits bone resorption and lowers the serum calcium.30 Given that the acute effect of CT on serum calcium is related to the prevailing rate of bone resorption, it is not surprising that CT has little or no effect on serum calcium in the mature animal or human subject, since the rate of bone resorption is slow in maturity. The physiologic function of CT in maturity nevertheless may be to regulate the bone resorptive process in a continuous or intermittent manner. It follows that CT should not necessarily be regarded as a “calcium-regulating hormone” in maturity, but may yet be shown to be such in stages of rapid growth (e.g., in the young, in states of increased bone turnover). It is nevertheless important that bone resorption be regulated, and CT is the only hormone known to be capable of carrying out this function through a direct action on bone. Physiologic roles for other members of the CT family, such as amylin and CGRP, in the regulation of resorption have not yet been substantiated. An antiresorptive role for CT in maturity might become more important in circumstances in which skeletal integrity is at particular risk (e.g., in pregnancy and lactation). Evidence in support of such an important physiologic role for endogenous CT in protecting against bone loss is provided by experiments showing that cancellous bone loss in thyroparathyroidectomized rats treated with PTH was greater than that in similarly treated sham-operated controls.31 In addition, mice in which the CT/CGRP gene was ablated showed a severe drop in bone mineral content during lactation, although the maternal skeleton recovered to baseline thereafter.32,33
CALCITONIN ACTIONS IN BONE
Osteoclasts
The first evidence of the mechanism of action of CT was obtained by showing in organ culture of bone that CT inhibited bone resorption.34 Inhibition of resorption appeared to be explained by a direct action on osteoclasts. CT treatment of resorbing bone in vitro resulted in rapid loss of osteoclast ruffled borders and decreased release of lysosomal enzymes. In vivo evidence was also consistent with an inhibitory action upon bone resorption. Loss of ruffled borders in osteoclasts was seen in patients with Paget’s disease, in whom bone biopsies were taken before and 30 minutes after an injection of CT.35 In the same clinical study, CT was noted to decrease the number of osteoclasts, in addition to altering their ultrastructure. CT infusion in rats led to an immediate reduction in the rate of excretion of hydroxyproline, consistent with inhibition of breakdown of bone collagen.36 Other studies led to similar conclusions, with no evidence to suggest any increase in the active uptake of calcium by bone.37
Studies of the actions of hormones on isolated bone cell populations established that CT acts directly on osteoclasts, with receptor autoradiography showing osteoclasts as the only discernible bone cell targets.38 Mammalian osteoclasts possess abundant, specific, high-affinity receptors for CT (Fig. 57-5), and CT stimulates cAMP formation in a sensitive and dose-dependent manner,38 as well as increasing intracellular free calcium levels and protein kinase C activity.39 As stated, the direct effect of CT upon the osteoclast was found to result in rapid inhibition of activity, reflected in cessation of motility and contraction of the cell. Although isolated osteoclasts remained quiescent in CT as long as the hormone was present, they regained activity when osteoblasts were added to the culture.40 This escape of osteoclasts from inhibition by CT took place at a rate proportional to the number of osteoblasts with which they were in contact. CT reduced the cytoplasmic spreading of isolated osteoclasts in a dose-dependent manner. PTH had no effect unless osteoblasts were co-cultivated with the osteoclasts, in which case addition of PTH resulted in a marked increase in cytoplasmic spreading of osteoclasts. It cannot be assumed that these phenomena reflect the responses of cells in bone in vivo, but this work provided for the first time some useful direct observations of actions of hormones on isolated bone cell preparations containing osteoclasts. These observations though may be relevant to our interpretation of recent findings in mice rendered null for the CT/CGRP gene and in those haploinsufficient for the CT receptor (vide infra).
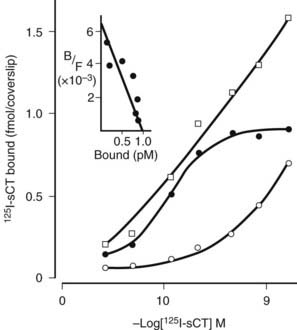
FIGURE 57-5. Saturation analysis of labeled calcitonin binding to rat osteoclasts. Scatchard analysis of specific binding (inset) shows receptor number of 4.8 × 106 per cell.
(Data from Nicholson GC et al, J Clin Invest 78:355–360, 1986.)
The molecular mechanisms by which CT decreases osteoclast function have yet to be fully defined. The rapid effects of the hormone may be brought about through actions on a cytoskeletal function of osteoclasts, after initial events involving generation of several intracellular second messengers. Early events in CT signal transduction have been studied in a variety of cell types and are described in greater detail later. The other means by which CT could inhibit resorption is through inhibition of osteoclast formation. In vivo data and results from CT inhibition of resorption in organ culture are suggestive of this. The development of methods of studying osteoclast formation in vitro from hemopoietic precursor cells has allowed this question to be addressed directly. Several reports have described CT inhibiting osteoclast-like cell formation in bone marrow cultures of human, baboon, and mouse origin.41–44 However, these experiments were all conducted at relatively high CT concentrations, and the effects were small. In other studies, in which lower concentrations of CT were used, which nevertheless reduced CT receptor mRNA expression in developing mouse osteoclasts, no reduction in osteoclast formation was observed.45–47 The multinucleated osteoclasts that formed in the continuous presence of exogenous CT had fewer nuclei though, and the osteoclasts generated under these conditions were deficient in CT receptor mRNA and protein but nevertheless capable of resorbing bone. In elegant studies of CT administration to mice, Ikegame et al48 showed that the CT-induced drop in serum calcium was linked temporally to the loss of osteoclast ruffled borders. Further, frequent dosing of the animals resulted in insensitivity to CT in terms of recovery of osteoclast ruffled borders and return of serum calcium to control levels. It was significant that treatment of mice with CT initially rendered osteoclasts unable to bind 125I-sCT, which recovered after a single treatment but not with repeated treatment.48 These findings may be relevant to the mechanism of “escape” from CT that is observed clinically.
Osteoblasts
Although the best understood action of CT in bone is as an antiresorptive agent, numerous reports have described actions also on cells of the osteoblast lineage, as well as direct and indirect effects on bone formation. CT increased [3H]thymidine incorporation in embryonic chicken calvariae, in the transformed murine calvarial cell lines MMB and MC-3T3-E1 and in primary cultures of cells prepared from newborn mouse calvaria.49 CT was also shown to stimulate [3H]thymidine incorporation in primary human osteoblasts50 and to increase expression of insulin-like growth factors (IGFs) by human SaOS-2 cells.51 None of these observations has been confirmed, and indeed data failing to show such effects of CT have been published.52 In addition to the fact that relatively high concentrations of CT were used in these early experiments, results with primary osteoblasts should be interpreted with caution at present, because it is known that calvarial osteoblast preparations are contaminated with osteoclast precursors, and a recent report provides evidence that primary osteoblast preparations contain a substantial proportion of bone-specific macrophages, which have been termed “osteomacs.”53 This raises the possibility that the presumed osteoblast responses may actually be mediated by cells of the monocyte-macrophage lineage. Furthermore, no published evidence is convincing of specific, functional CT receptors in any cells of the osteoblast lineage. Inadequate criteria were used in a claim that 125I-sCT was bound specifically to osteocyte-like cells, MLO-Y4.54 In the same work, CT at high concentrations was associated with a small increase in cAMP and with protection of the cells from apoptosis induced by etoposide, tumor necrosis factor (TNF)-α or dexamethasone. It has been claimed that CT can hasten and improve the process of fracture healing in normal55 and osteoporotic rats.56 If this proves to be the case, the mechanism could be similar to that of bisphosphonates, which have also been shown to enhance the strength of the healed fracture, apparently by modulating resorption during the remodeling phase of bone repair.57 It could also relate to possible stimulation of angiogenesis by CT, which has been shown for human microvascular endothelial cells, albeit at supraphysiologic concentrations.58
NEW UNDERSTANDING OF THE ROLE OF CALCITONIN IN BONE: STUDIES IN GENETICALLY MANIPULATED MICE
The preceding discussion of the physiology of CT and its action on bone reflects views that have remained largely unaltered over many years, with few new data to change them. The data on which they are based, particularly since normal and low circulating CT levels cannot be measured with confidence (vide supra), do not provide a convincing argument for a specific physiologic role for calcitonin. Indeed, in a recent review, one of the co-discoverers of calcitonin59 argued the case that calcitonin is not involved in calcium homeostasis or in any other important physiologic function, except possibly in protection of the skeleton under conditions of calcium stress. Some recent work has changed this situation. Ablation of the CT/CGRP gene in mice resulted in viable and fertile mice with no production of CT or CGRPα.32 As expected, these mice were much less able than wild-type mice to overcome the hypercalcemia induced by a calcium load,32 and they lost excessive bone during lactation.33 The great surprise with these mice, however, was that they had increased bone mass, with histomorphometric parameters showing increased bone formation.32 This suggested that a normal role for calcitonin might be that of an inhibitor of bone formation. The finding was unexpected and counterintuitive, and it was possible that the dual ablation of CT and CGRPα might explain it, although there was no obvious mechanism for this. Indeed, the increased bone formation phenotype was not found when CGRPα-deficient mice were examined,60 thereby making calcitonin deficiency likely responsible for the increased bone formation. In reviewing the role of calcitonin, Hirsch59 had considered the results obtained with the CT/CGRP−/− mice but regarded them as inconclusive; as information accumulates, the new physiology of calcitonin is becoming more apparent. The CT/CGRP-deficient mice were protected against ovariectomy-induced bone loss,32,60 and, it was striking that after the age of 6 months these mice showed severe cortical porosity, even though indices of increased bone formation were maintained.61,62 Taken together, the observations are indicative of an inhibitory effect of calcitonin on bone formation, most likely an indirect one, and a direct effect on bone resorption through action on the osteoclast.
The significance and importance of these findings were enhanced however with the outcome of the studies of Dacquin et al63 in mice in which the CT receptor (CTR) was genetically manipulated. CTR−/− mice were embryologically lethal, and this was thought to be due to a placental effect. The CTR+/− mice, however, exhibited a bone phenotype virtually indistinguishable from the CT/CGRP−/− mice, with increased bone mass and increased bone formation on histomorphometry. Thus, the conclusion is that in mice the removal of calcitonin production or action results in an increased amount of bone, implying a physiologic role for calcitonin as a tonic inhibitor of bone formation. The same group prepared mice at least 94% deficient in the CTR and again found evidence of increased indices of bone formation.64 How the effect on bone formation comes about remains to be determined. The lack of evidence for specific calcitonin receptors and responses in osteoblasts is compelling, so it is very likely that the physiologic roles of calcitonin in bone are brought about through two pathways: a direct effect on osteoclasts to inhibit resorption and an indirect one resulting in the elaboration of a critical, locally active factor that is necessary for bone formation. This could result from signals through receptors in the osteoclast or in the hypothalamus.62,65 Resolution of this question will be a matter of very great interest, especially in light of ample recent evidence for central regulation of bone metabolism.66
RENAL ACTIONS OF CALCITONIN
When infused into thyroparathyroidectomized rats, CT caused a dose-dependent phosphaturia, but the effect on phosphate excretion was only a minor one in comparison with the phosphaturic effect of PTH.67 Although this was demonstrated in human subjects also, in several species CT failed to have any effect on phosphate excretion. Thus, it seems unlikely that the phosphaturic effect is of any major physiologic significance.
A number of other renal effects of CT, including a transient increase in calcium excretion due probably to inhibition of renal tubular calcium reabsorption, have been noted.68 Although this has not usually been regarded as an important effect of CT, it has been linked to the calcium-lowering effect of CT in hypercalcemic patients with metastatic bone disease. The use of CT in the treatment of hypercalcemia due to cancer has been based exclusively on the inhibition of osteolysis by CT. Some evidence has been produced that failure of the kidneys to excrete the calcium load derived from bone breakdown is a major contributor to the hypercalcemia. This has prompted careful study of the relative contributions to the hypocalcemic effect of CT of its renal and skeletal components. It was concluded that inhibition of renal tubular reabsorption by CT can induce a rapid fall in serum calcium, and that the magnitude of this effect depends upon the correction of volume depletion, which inevitably accompanies hypercalcemia.69 Thus, the calciuretic action of CT may assume greater importance than was hitherto suspected.
CT receptors are present in rat kidney,70 and the action of CT upon adenylate cyclase activity has been localized in the human nephron, predominantly to the medullary and cortical portions of the thick ascending limb and to the early portion of the distal convoluted tubule. The co-localization of the CT receptor mRNA expression and cell surface receptors with G protein–sensitive adenylate cyclase is consistent with cAMP being an important mediator of CT action in this organ. A possible role for CT in the kidney is to regulate 1,25(OH)2D3 levels, with an original observation of enhanced 1-hydroxylation of 25(OH)D in the proximal straight tubule of the kidney by CT stimulation of the expression of 25(OH)D 1α-hydroxylase.71 Subsequently, Shinki et al72 showed that CT administration to rats induced renal CYP27B1 when serum calcium levels were normal or high, and this was supported by a report that CT treatment in rats increased renal production of CYP27B1 mRNA.73 CT stimulation of the expression of CYP24 in CTR-transfected HEK-293 cells has also been reported.74 The authors speculated that, since 1,25(OH)2D3 and CT synergistically stimulate CYP24 mRNA production in kidney cells, this latter action of CT could be part of the process by which it regulates serum calcium by controlling renal production of 1,25(OH)2D3.
Peptides Related to Calcitonin
The CT peptides are homologous with the related peptides CGRP, amylin, and the adrenomedullins. Greatest identity is between the teleost CTs and amylin (≈33%), with ≈22% identity with the CGRPs and 16% with adrenomedullin. Less homology is observed between the mammalian CTs and the other peptides. Consistent with the homology between peptides, there is a limited degree of overlap in specificity among the binding sites for the peptide receptors, with CT-like actions seen with high concentrations of amylin and CGRP.
Amylin is a 37 amino acid peptide that is co-secreted with insulin from pancreatic β cells following nutrient ingestion. Amylin at physiologic concentrations is important in the integrated control of nutrient influx with potent actions, including inhibition of gastric emptying, gastric acid secretion, food intake, digestive enzyme secretion, and glucagon secretion.75 Amylin at higher concentrations also acts to inhibit insulin secretion from the pancreas and to promote glycogen breakdown and to decrease insulin-stimulated incorporation of glucose into glycogen in skeletal muscle. Thus, amylin is thought to act as a partner to insulin in metabolic regulation, although this effect may not occur at normal circulating levels of the peptide.76 In the kidney, amylin is proposed to have a diverse range of actions, including modulation of Ca2+ excretion and thiazide receptor levels, proliferative effects on tubule epithelium, and increasing renin activity.77,78 Amylin−/− mice have been shown to have less bone as the result of increased bone resorption, and in vitro tests indicated that this may be due to release of an amylin-mediated attenuator of osteoclastogenesis.63 These authors speculate that the receptor for this effect is independent of the CTR gene, which, with receptor activity modifying proteins (RAMPs), forms the basis of characterized amylin receptor phenotypes (see later). However, their conclusion is based on studies in animals with the amylin−/+, CTR−/+ genotype, and therefore RAMP/CTR-based amylin receptors cannot be excluded as the target for amylin action.
Amylin receptors are also widely expressed in brain, where administered peptide induces many potent effects. These include decreased appetite and gastric acid secretion, hyperthermia, adipsia, and reduction in growth hormone–releasing hormone. Central amylin injection may also modulate memory and the extrapyramidal motor system.75,77 The molecular basis for amylin receptor phenotype is discussed later.
CGRP is a pleiotropic neuropeptide with a diverse range of actions including potent dilation of vascular beds, as well as relaxation of other smooth muscle, inotropy and chronotropy in the heart, and paracrine regulation of pituitary hormone release, and many central effects, such as suppression of appetite and gastric acid secretion, modulation of body temperature, and modulation of sympathetic outflow. CGRP also acts to modulate nicotinic acetylcholine receptor levels at neuromuscular junctions. CGRP weakly modulates calcium homeostasis, although this is likely to reflect its low affinity for interaction with CTRs. The actions of CGRP have been extensively reviewed elsewhere.78,79 Specific CGRP receptors have been characterized in many tissues and it is likely that more than one subtype of receptor exists. CGRP receptors arise from hetero-oligomerization of RAMP1 with either the calcitonin receptor-like receptor (CLR) or CTR,80,81 although weaker interactions are also seen with other RAMP/CLR- or RAMP/CTR-based receptors.82
Adrenomedullin was originally isolated from human pheochromocytoma and is abundant in the normal adrenal medulla, hence its name. The full-length peptide of ≈50 amino acids shares approximately 25% homology with CGRP across its N-terminal 37 amino acids.83 Adrenomedullin is a potent dilator of many vascular beds and is protective against conditions such as cardiac hypertrophy, perivascular fibrosis, renal damage, and pulmonary hypertension.84,85 Adrenomedullin receptors arise from heterodimers of CLR and RAMP2 or RAMP3.81 Both adrenomedullin and amylin can stimulate osteoblast proliferation at low concentration.86 Fig. 57-6 illustrates complex formations between the CTR and RAMPs 1, 2, and 3.
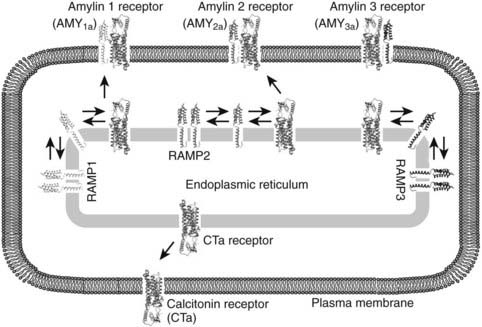
FIGURE 57-6. Representative illustration of complex formation between the calcitonin receptor (CT) and RAMPs 1, 2, and 3 in the endoplasmic recticulum (ER). All three RAMPs are postulated to exist as homodimers in the ER; however, in the presence of CTR an equilibrium is established in which hetero-oligomers between the receptor and each of the RAMPs are formed. To date, the stoichiometry of the oligomers remains unclear. AMY1a, AMY2a, and AMY3a receptors are generated when the CTa receptor isoform is complexed with RAMP 1, 2, and 3, respectively. In addition, although the oligomers between the CT receptor and RAMPs can then be transported to the cell surface, the CT receptor is transported to the cell surface in the absence of RAMPs. The human CT receptor has two major splice variants, CTa and CTb, that arise from the absence or presence of a 16 amino acid insert in intracellular domain 1. The nomenclature for amylin receptors formed by the different receptor isoforms is denoted by (a) or (b) in the name (e.g., AMY1a denotes an amylin receptor formed from the CTa receptor and RAMP 1). It has been hypothesized that RAMPs may reside in a homodimerized form in intracellular sorting compartments such as the endoplasmic reticulum.
Recently, new peptides related to the CT family of peptides have been identified. The calcitonin receptor–stimulating peptides (CRSPs) were originally isolated/cloned from porcine brain and, being localized principally to brain and pituitary, have been proposed as potential endogenous ligands for central CTRs.87–89 However, not all CRSPs activate CTRs or related receptors, suggesting that they may also stimulate other receptors.88 To date, homologous peptides have not been identified in humans or rodents, although bovine and canine homologues do exist.89 In addition, a second adrenomedullin-like peptide (adrenomedullin 2; also known as intermedin) has been identified in multiple species, including human. This peptide has similar affinity for both CGRP and adrenomedullin receptors, although the physiologic significance of this peptide is still unclear.82,90,91
Calcitonin in the Central Nervous System
Intraventricular administration of CT generates potent effects that include analgesia and inhibition of appetite and gastric acid secretion, as well as modulation of hormone secretion and the extrapyramidal motor system.77,79,81,92 Both immunoreactive CT-like peptide and CT receptors have been demonstrated in the brain and nervous system of rats, humans, and other species. Immunoreactive CT related antigenically to hCT (hCTI) has also been demonstrated in the nervous systems of protochordates, lizards, and pigeons, as well as at low levels in human and rat brain extracts. Furthermore, radioimmunoassay analyses point to the presence of sCT-like peptide material in human and rat brain.93
Low levels of hCTI occur in extracts of postmortem human brain. However, in addition to hCTI, low levels of material chromatographically and immunologically similar to sCT (sCTI) have been found, with concentrations in the brain ≈10 times greater than those in serum and cerebrospinal fluid. Similarly, extracts of rat brain diencephalon contain an sCT-like peptide.93 Although hCTI has been detected in rat brain, only limited evidence has been found for the presence of rat CT (rCT) mRNA. Immunoreactive CT-like material also occurs in the pituitary of both mammals and lower vertebrates, although the identity of the pituitary CT remains to be established. The physiologic significance of CT-like immunoreactivity in the pituitary remains to be fully elucidated; however, CT receptors are present in the intermediate pituitary, and therefore the CT-like material may act as a paracrine regulator of these receptors. CT has been recently described as participating in a complex cross-talk between extracellular signaling molecules in the pituitary.94 Treatment of pituitary cells with CT in culture modulates the secretion of prolactin,95,96 and treatment of the cultures with an anticalcitonin antibody enhances it.97 Targeted overexpression of calcitonin in gonadotrophs of mice leads to long-term hypoprolactinemia, decreased PRL gene expression, female subfertility, and a selective underdevelopment of lactotrophs.98,99
Related posts:






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


