The dense tubular system is also the site of prostaglandin (PG) synthesis in platelets.26,27
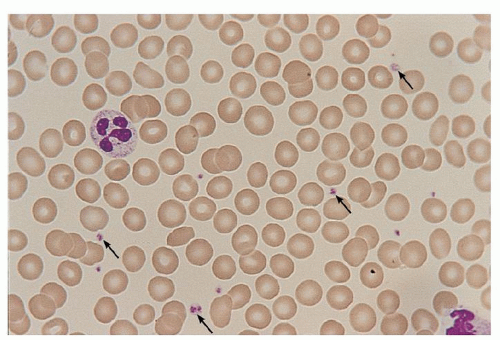 FIGURE 16.1. A human peripheral blood smear stained with Wright-Giemsa. Platelets, indicated by arrows, are interspersed between erythrocytes and a few leukocytes. The pale, grayish-blue cytoplasm contains purple-red granules. Original magnification of 35-mm slide ×100. |
 FIGURE 16.2. Scanning electron micrograph of unstimulated human platelets. Most are discoid (d) in shape. Many surface indentations, indicated by arrows, are present; these correspond to openings of the surface-connected canalicular system to the external milieu. Magnification ×15,000. (Data from Stenberg PE, Shuman MA, Levine SP, Bainton DF. Optimal techniques for the immunocytochemical demonstration of β-thromboglobulin, platelet factor 4, and fibrinogen in the alpha granules of unstimulated platelets. Histochem J 1984;16:983-1001.) |
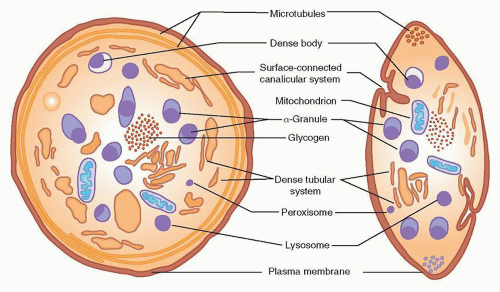 FIGURE 16.3. Diagram of a human platelet displaying components visible by electron microscopy and cytochemistry. In addition to membranous components (plasma membrane, surface-connected canalicular system, and dense tubular system), mitochondria, microtubules, and glycogen, four types of storage organelles are identified: α-granules, dense bodies, lysosomes, and microperoxisomes. Whereas the first two can be identified morphologically, microperoxisomes and lysosomes are recognizable only by cytochemical stains. (From Bentfeld-Barker ME, Bainton DF. Identification of primary lysosomes in human megakaryocytes and platelets. Blood 1982;59:472-481, with permission.) |
rise in intracellular calcium, the proportion of F-actin increases rapidly to 60% or 70%.46 Actin monomers polymerize into filaments at platelet peripheries,45,47 and bundles of new filaments form to fill developing filopodia.48,49 and 50 Phosphorylation of myosin light chain results in binding of myosin to actin,51,52,53 providing the tension required for granule centralization and retraction of filopodia.54 Additional proteins, such as talin and surface αIIbβ3, join the developing electron-dense actin filaments, and the structure is remodeled through the action of an associated calciumdependent protease, calpain.55 The sum of these events in platelet function is critical because the combination of various additions, rearrangements, and remodeling steps underpins shape change and spreading (filopodia and lamellipodia formation), along with platelet secretion and clot retraction.
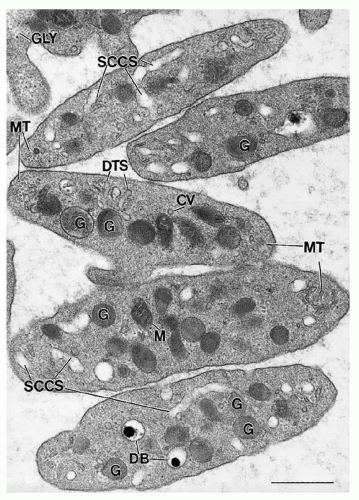 FIGURE 16.4. Ultrastructure of unstimulated human platelets. Membranous organelles, including the surface-connected canalicular system (SCCS) and dense tubular system (DTS), and cytoplasmic organelles, including mitochondria (M), α-granules (G), dense bodies (DB), coated vesicles (CV), and glycogen (GLY), are visualized at the ultrastructural level. Microtubules (MT) are present as cross-sectional and longitudinal profiles at the poles of the discoid platelets. Magnification ×46,000. Bar = 0.5 µm. |
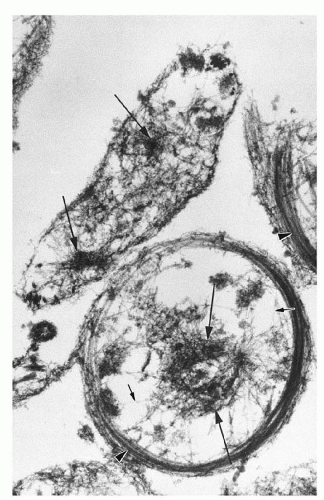 FIGURE 16.5. Human platelet cytoskeletons prepared by simultaneous fixation and lysis in Triton X-100 detergent. Single actin filaments, indicated by short arrows, course throughout the platelet cytoplasm. Clusters of filaments, indicated by long arrows, also are present. Note the microtubule coils at the platelet peripheries (arrowheads). Magnification ×30,000. (From Boyles J, Fox JEB, Phillips DR, Stenberg PE. Organization of the cytoskeleton in resting, discoid platelets: preservation of actin filaments by a modified fixation that prevents osmium damage. J Cell Biol 1985;101:1463-1472, with permission.) |
along with factor V.65,66,67 Platelets take up plasma proteins and store them in their α-granules.68,69,70 Select α-granule proteins are discussed below and listed in Table 16.2.
TABLE 16.1 MAJOR PLATELET CYTOSKELETAL PROTEINS | |||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||
sulfatase, β-hexosaminidase, β-galactosidase, heparitinase, and β-glycerophosphatase. Additional proteins found in lysosomes include cathepsin D and lysosome-associated membrane proteins (LAMP-1/LAMP-2), which are expressed on the plasma membrane after activation).106,107 Lysosomal constituents are released more slowly and incompletely (maximally, 60% of the granules) than α-granules or dense-body components after platelet stimulation, and their release also requires stronger agonists such as thrombin or collagen.
TABLE 16.2 MAJOR PLATELET GRANULAR CONSTITUENTS SECRETED WITH ACTIVATION | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
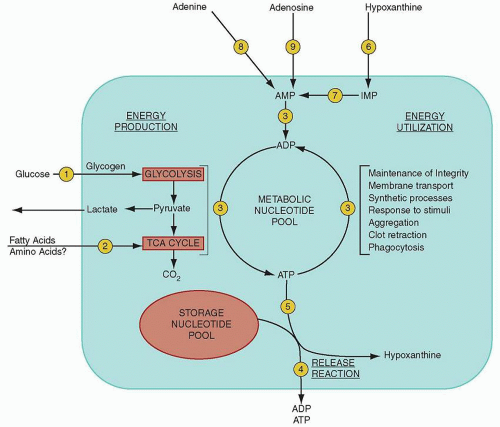 FIGURE 16.6. Simplified scheme of platelet energy metabolism. Platelet energy is derived from the metabolism of glucose and, to a lesser extent, from the metabolism of fatty acids. Energy is provided in approximately equal amounts by glycolysis and the citric acid cycle. The platelet energy reserve is provided by the metabolic pool of platelet nucleotides that is in a state of continuous turnover. This energy is used for the maintenance of the platelets’ structural integrity and in the reactions accompanying the response of platelets to stimuli. The granule-bound storage (nonmetabolic) nucleotide pool is discharged during the release reaction. ADP, adenosine diphosphate; AMP, adenosine monophosphate; ATP, adenosine triphosphate; IMP, inosine monophosphate. (Adapted from Hirsh J, Doery JCG. Platelet function in health and disease. Prog Hematol 1972;7:185-234.) |
cytoplasmic pool makes up 40% of total adenine nucleotides; it is used for the maintenance of various energy-consuming cell functions and is retained during platelet release. In large part made up of ATP, this pool is constantly turning over, as revealed by the rapid incorporation of 14C-adenine and 32P-phosphate into ATP. In unstimulated platelets, the relative concentrations of metabolic AMP, ADP, and ATP are maintained by the enzyme adenylate kinase (Fig. 16.6, Step 3).128
vigorous exercise, and this rise is not affected by splenectomy.165 This nonsplenic pool represents approximately 16% of the total platelet mass.
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


