that are upregulated to the cell surface with stimulation.2 The relative contribution of tertiary granules and specific granules to upregulation of membrane proteins is not clear.
TABLE 7.1 CONTENTS OF HUMAN NEUTROPHIL GRANULES | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
TABLE 7.2 SOME CD ANTIGENS EXPRESSED ON NEUTROPHILS | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
evidence that in neutrophils, proteins may enter these rafts when they are translocated to the cell surface. For example, it appears that CD63 and CD11b/CD18 are not present in detergent-resistant complexes when they are intracellular, but they enter such complexes after translocation to the cell surface.63
TABLE 7.3 MORPHOLOGIC CHARACTERISTICS OF LEUKOCYTES (WRIGHT STAIN) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
all subsequent stages, including the polymorphonuclear forms (Fig. 7.1).97,103,104
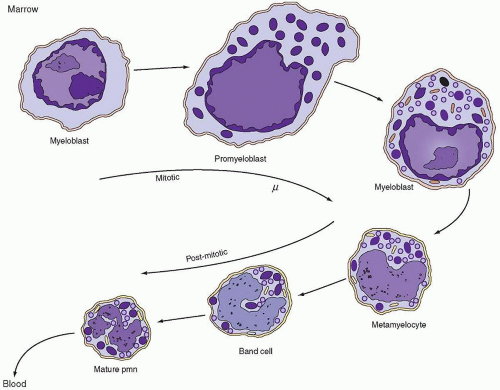 FIGURE 7.1. Appearance of granules during neutrophil maturation. Myeloblasts are undifferentiated cells with a large oval nucleus, large nucleoli, and cytoplasm lacking granules. They originate from a precursor pool of stem cells. Subsequently, there are two stages—the promyelocyte and the myelocyte—each of which produces a distinct type of secretory granule: azurophils (dark granules) are produced only during the promyelocyte stage; specific granules (light granules) are produced during the myelocyte stage. The metamyelocyte and band forms are nonproliferating stages that develop into the mature polymorphonuclear neutrophil characterized by a multilobulated nucleus and cytoplasm containing primarily glycogen and granules. Both nonspecific azurophilic granules and specific granules persist throughout these later stages. PMN, polymorphonuclear leukocyte. (Modified from Bainton DF, Ullyot JL, Farquhar MG. The development of neutrophilic PMN leukocytes in human bone marrow: origin and content of azurophil and specific granules. J Exp Med 1971;134:907.) |
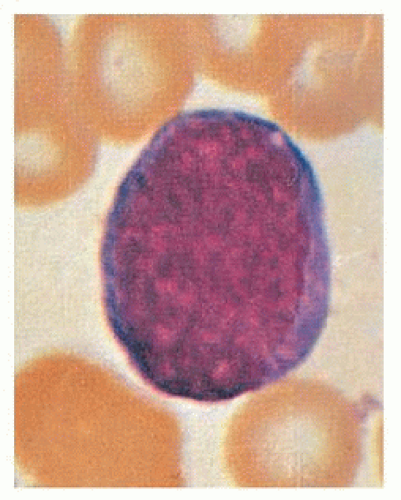 FIGURE 7.2. Myeloblast (×1,000, Wright stain). |
persist in myelocytes, but formation of new primary granules is limited to the promyelocyte, and each succeeding cell division leads to a decrease in their number in the daughter population (Fig. 7.1).97,104,105 The secondary granules of the neutrophil series are smaller than the primary granules; in the rabbit, cat, and human, they are formed in increasing numbers on the convex surface and lateral borders of the somewhat less prominent Golgi apparatus.97,102,104 The amount of granular endoplasmic reticulum is lower in the myelocyte than in earlier forms, so the cytoplasmic basophilia decreases and disappears. The mitochondria remain small and are few.
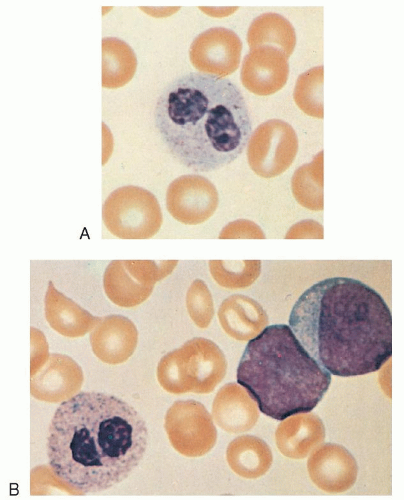 FIGURE 7.3. A, B: Pseudo-Pelger-Huët cells, the latter from the blood of a patient with acute myeloblastic leukemia (×1,000, Wright stain). |
have regarded a constriction greater than one half or two thirds of the nuclear breadth as adequate evidence of lobulation and classify such cells as polymorphonuclear128,129 or use slightly different criteria.130 Still others avoid the issue entirely. Because no clear difference has been shown between band and segmented stages other than nuclear shape and a slightly earlier appearance of 3H-TdR in the band forms, the distinction becomes arbitrary. However, a clear and easily recognizable separation is needed if one wishes to count nuclear lobes for diagnostic purposes, as in the early detection of folic acid deficiency131 or in assessing marrow release of young forms into the blood.132 For such purposes, we have chosen the clear separation of nuclear lobes as the criterion for inclusion in the polymorphonuclear category.127 Cells without this complete formation of distinct lobes (usually connected by a filamentous strand) are classified as band forms.
 FIGURE 7.4. Early and late promyelocytes, a myelocyte, and a polymorphonuclear neutrophil (PMN) viewed by electron microscopy. (Courtesy of Bainton D, University of California, San Francisco.) A: Early neutrophilic promyelocyte (reacted for peroxidase, 10,500). The nucleus (N) with its prominent nucleolus (Nu) occupies the bulk of this immature cell. The surrounding cytoplasm contains a few azurophil granules (ag), a large Golgi complex (G), Golgi cisternae (Gc), several mitochondria (µ), scanty rough endoplasmic reticulum (er), and many free polysomes (r). A centriole (ce) is present in the Golgi region. All of the azurophil granules (ag) appear dense because they are strongly reactive for peroxidase. The secretory apparatus, i.e., the perinuclear cisterna (pn), rough endoplasmic reticulum (er), and Golgi cisternae (Gc), are also reactive, although less so than the granules. Specimen was fixed in glutaraldehyde for 16 hours at 4°C, incubated in the peroxidase medium of Graham and Karnovsky for 1 hour at 22°C, postfixed in osmium tetroxide, treated in block with uranyl acetate, dehydrated in ethanol, infiltrated with propylene oxide, and embedded in Araldite. Section stained for 1 minute with lead citrate. B: Late neutrophilic promyelocyte (reacted for peroxidase, 7,000). This cell is the largest (15 µm) of the neutrophilic series. It has a sizable, slightly indented nucleus (N), a prominent Golgi region (G), and cytoplasm packed with peroxidase-positive azurophil granules (ag). Note the two general shapes of the azurophil granules: spherical (ag) and ellipsoid (ag′). Most are spherical, with a homogeneous matrix, but a few ellipsoid forms containing crystalloids also are present. Many of the spherical forms (ag) have a dense periphery and a lighter core, presumably because of incomplete penetration of substrate into the compact centers of mature granules. Peroxidase reaction product is visible (under higher magnification) in less concentrated form within all compartments of the secretory apparatus (endoplasmic reticulum, perinuclear cisterna, and Golgi cisternae). No reaction product is seen in the cytoplasmic matrix, mitochondria, or nucleus. Specimen was fixed in glutaraldehyde for 10 minutes at 4°C and subsequently processed exactly as was the specimen in A. C: Neutrophilic myelocyte (reacted for peroxidase, 9,000). At this stage, the cell is smaller (10 µm) than the promyelocyte, the nucleus is more indented, and the cytoplasm contains two different types of granules: large, peroxidase-positive azurophils (ag) and the generally smaller specific granules (sg), which do not stain for peroxidase. A number of immature specific granules (is), which are larger, less compact, and more irregular in contour than mature granules, are seen in the Golgi region (G). Note that peroxidase reaction product is present only in azurophil granules and is not seen in the rough endoplasmic reticulum (er), perinuclear cisterna (pn), and Golgi cisternae (Gc), in keeping with the fact that azurophil production has ceased, and only peroxidase-negative specific granules are produced during the myelocyte stage. D: Mature PMN (reacted for peroxidase, 10,500). The cytoplasm is filled with granules; the smaller peroxidase-negative specific granules (sg) are more numerous, the azurophils (ag) having been reduced in number by cell divisions after the promyelocyte stage. Some small, irregularly shaped azurophil granule variants are also present (unlabeled arrow). The nucleus is condensed and lobulated (N1-N4), the Golgi region (G) is small and lacks forming granules, the endoplasmic reticulum (er) is scanty, and mitochondria (µ) are few. Note that the cytoplasm of this cell has a rather ragged, moth-eaten appearance because the glycogen, which is normally present, has been extracted in this preparation by staining in block with uranyl acetate. |
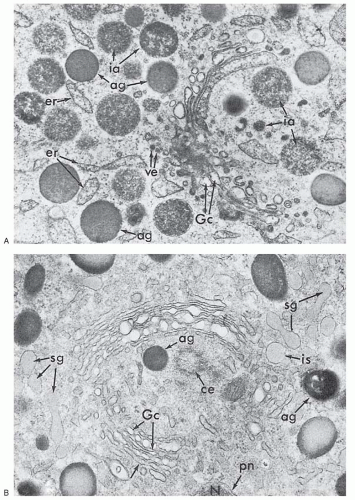 FIGURE 7.5. Granule formation in neutrophil precursors viewed by electron microscopy. (Courtesy of Bainton D, University of California, San Francisco.) A: Golgi region of a neutrophilic promyelocyte reacted for peroxidase (×40,000). At this stage, the peroxidase reaction product is present within the rough endoplasmic reticulum (er), the clusters of smooth vesicles (ve) at the periphery of the Golgi cisternae (Gc), in the Golgi cisternae, and in the immature (ia) and mature (ag) azurophilic granules. The immature granules are larger and less compact than the uniformly dense mature granules. B: Golgi region of a neutrophilic myelocyte reacted for peroxidase (×40,000). Peroxidase-reactive material is seen in the primary or azurophilic granules (ag) but not in the specific (secondary) granules (sg). At this stage (myelocyte), no peroxidase reaction product is seen in the endoplasmic reticulum, Golgi cisternae (Gc), or newly formed, immature specific granules (is). The stacked Golgi cisternae are oriented around the centriole (ce), and the outer cisternae (unlabeled arrow) contain material of intermediate density that is similar to the content of the specific granules, suggesting that the specific granules arise from the convex face of the Golgi complex as in the rabbit. pn, perinuclear cisternae. |
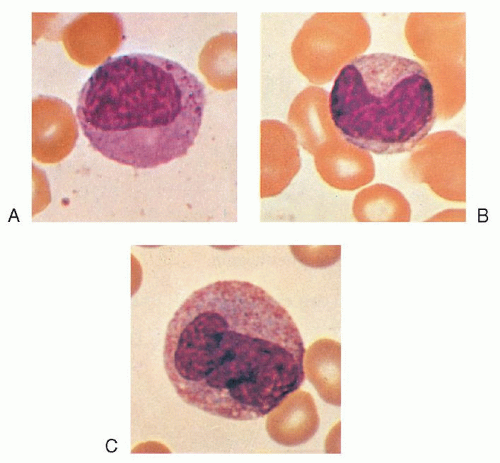 FIGURE 7.6. A: Late myelocyte or early metamyelocyte. B: Metamyelocyte. C: Monocyte (×1,000, Wright stain). |
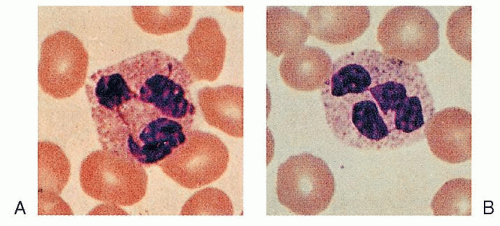 FIGURE 7.7. A, B: Polymorphonuclear neutrophils. |
Neutrophil heterogeneity has also been demonstrated in the case of olfactomedin 4 expression. Olfactomedin 4 is highly induced in myeloid progenitors by G-CSF. Although olfactomedin 4 mRNA is expressed in all myelocytes and metamyelocytes, only ˜25% of neutrophils in peripheral blood contain olfactomedin 4, which is localized to the specific granules.150
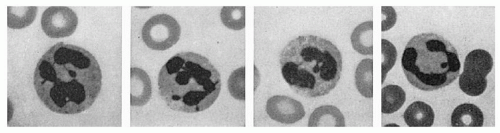 FIGURE 7.8. Granulocytes and sex chromatin patterns. Two cells on the left show the characteristic drumsticks found in the female subjects. The thin strand of chromatin joining the head to a nuclear lobe can be seen clearly. In the two cells on the right, small clubs, such as may be seen in male subjects, should not be confused with drumsticks (Wright stain, ×1,300). |
and black Africans, who have significantly lower neutrophil and monocyte counts.165 Lower counts are less evident in black Americans127 and in Africans eating a Western diet.163
TABLE 7.4 NORMAL BLOOD LEUKOCYTE CONCENTRATIONS (95% CONFIDENCE LIMITS) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
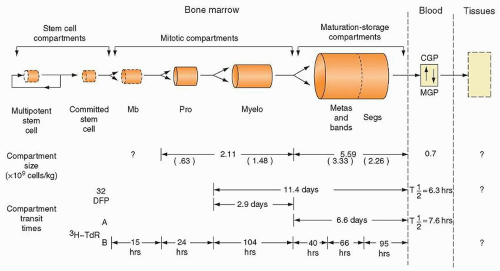 FIGURE 7.9. Model of the production and kinetics of neutrophils in humans. The marrow (555) and blood compartments (147) are drawn to show their relative sizes. In the lower one third of the figure, the compartment transit times as derived from di-isopropylfluorophosphate (DF32P) studies (138,147) and from tritiated thymidine (3H-TdR) studies (151,555) are compared. CGP, circulating granulocyte pool; MB, myeloblast; MGP, marginal granulocyte pool; myelo, myelocyte; pro, promyelocyte. |
throughout several half-times are needed, and that grain counting is extraordinarily tedious and subject to considerable error. Nevertheless, estimates of compartment turnover time have been made with this method by using 3H-TdR and radiosulfate.122
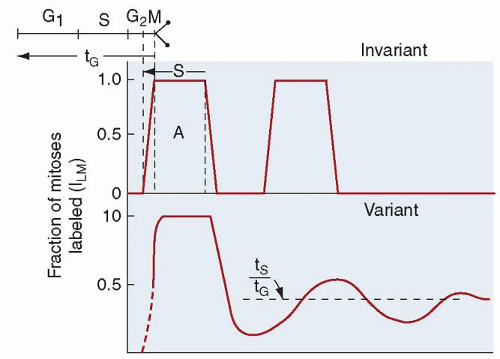 FIGURE 7.10. The percentage of labeled mitoses in the course of cell generation. At the top of the diagram is the theoretical configuration that would be seen if cells flowed through the proliferation cycle with no variation. The effects of moderate variation in time spent in the several cycle phases on the percent labeled mitosis curve are shown at the bottom. G1 is the pre-DNA synthesis resting phase (gap), S is the DNA synthetic period, G2 is the post-DNA synthesis gap, and M is mitosis. tS is the time spent in S, and tG is the duration of the entire generative cycle. (From Cronkite EP, the Brookhaven National Laboratory, and the National Cancer Institute. Kinetics of granulocytopoiesis. Natl Cancer Inst Monogr 30, Human Tumor Cell Kinetics 1969;51.) |
Related posts:
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


