tissues, both normally and in a range of inflammatory, infectious, metabolic, and neoplastic diseases.
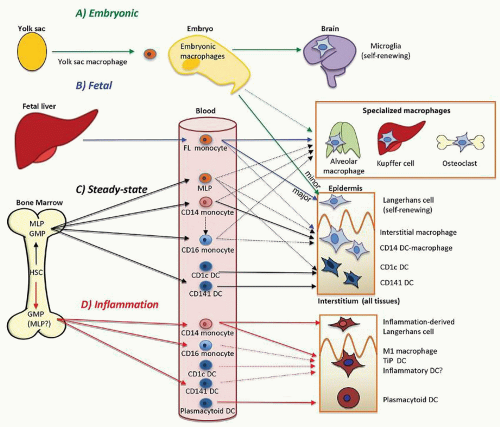 FIGURE 10.1. Summary of the main pathways of monocyte, macrophage, and dendritic cell (DC) development in human tissue during embryogenesis, fetal development, adult steady state, and inflammation. A: Embryonic. Yolk sac macrophages are the first hematopoietic cells to appear. Studies in mice indicate that these cells migrate through the mesoderm prior to vasculogenesis, proliferating and colonizing all the tissues of the embryo. Lineage tracing indicates that 100% of microglia arise from the yolk sac, becoming isolated in the developing brain by closure of the blood-brain barrier. They are maintained in adult life by local proliferation. B: Fetal. In models derived from lineage tracing in mice, hematopoiesis in the liver gives rise to the first monocytes, which circulate widely and dilute the contribution of yolk sac macrophages in almost all tissue sites. Residual yolk sac contribution of approximately 10% is detectable in the Langerhans cells of the epidermis; the other 90% are fetal liver-derived and become locally self-renewing in adult life. Tissue macrophages and specialized populations of macrophages: alveolar, Kupffer, and osteoclasts, may also derive from fetal liver monocytes, at least initially. C: Adult steady-state hematopoiesis, arising from multi-lymphoid progenitors (MLP) capable of differentiating into monocytes, DCs, lymphoid cells, and granulocyte-macrophage progenitors (GMP), contributes a wide variety of cells with macrophage and DC potential into the blood. These include circulating CD34+ MLP, classical CD14+ and nonclassical CD16+ monocytes, CD1c+ myeloid DCs, CD141+ myeloid DCs, and plasmacytoid DCs. The contribution of these steady state blood components to macrophage homeostasis in a wide range of tissues, in addition to those illustrated, has not been rigorously defined. CD34+ precursors and both subsets of monocytes potentially contribute to tissue macrophages in interstitial and specialized sites. CD14+ monocytes and CD16+ monocytes are closely related to CD14+ interstitial DCs, while CD1c+ and CD141+ blood DCs both have phenotypically similar interstitial DC counterparts. CD34+ precursors also have the potential to form tissue DCs. Plasmacytoid DCs do not migrate into the tissues in the steady state. D: Inflammation. Upregulation of myelopoiesis under conditions of stress is likely to involve expansion of GMP production of monocytes, myeloid DCs, and plasmacytoid DCs. Multiple inflammatory signals potentially recruit all peripheral blood populations to the tissues. Classical CD14+ monocytes are the most significant component giving rise to inflammatory activated “M1” macrophages and TNF/i-NOS-producing DCs (Tip-DCs) in a range of tissues. This may include the CNS (pathway not shown for simplicity here). Langerhans cells are also replaced by inflammatory classical monocytes in desquamating epidermal inflammation. Myeloid DCs may be directly recruited to give inflammatory DCs, and infiltration with plasmacytoid DCs is well documented. |
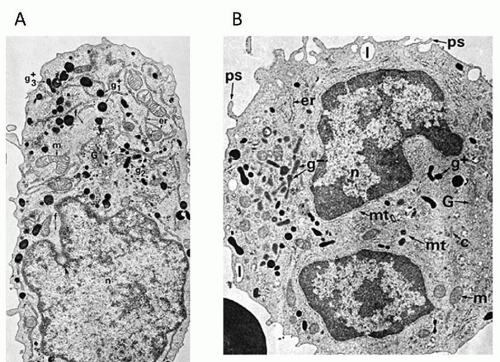 FIGURE 10.2. Ultrastructure of promonocyte and monocyte. A: Electron micrograph of a promonocyte from human bone marrow stained for peroxidase. The nucleus (n), situated at one end of the cell, exhibits an irregular outline and deep indentation. The cytoplasm contains a number of cytoplasmic organelles. Peroxidase reactivity is demonstrable throughout the rough endoplasmic reticulum (er), Golgi complex (G), and all cytoplasmic granules (g+1, g+2, g+3). Apparently, all granules mature from the earliest forms, which are spherical and dense (g+1), with a homogenous matrix, to more condensed and elongated forms (g+2), and then to dumbbell forms (g+3). The Golgi complex (G) is composed of several stacks of cisternae and occupies a large area adjacent to the nucleus. Bundles of filaments (f) are prominent in the cytoplasm and are believed to be useful in characterizing the cell as a monocyte form. Several mitochondria (m) are also seen ×18,000. B: Electron micrograph of a normal human monocyte examined for peroxidase. In the nucleus (n), the chromatin is more condensed than in earlier forms, is mainly peripheral in distribution, and is interrupted at the nuclear pores. The voluminous cytoplasm (c) contains a full complement of organelles associated with protein synthesis and export of secretory granules. Peroxidase is present in only some of the granules (g+), but others (g:), as well as the endoplasmic reticulum (er) and Golgi complex (G), now lack the reaction product. At this stage, the two kinds of granules are approximately equal in number and similar in size and shape, ranging from 90 to 450 nm in length and from spherical or rodlike to dumbbell in shape. Microtubules (mt) radiate from the cell center, where a centriole can be seen adjacent to the Golgi complex (G). The moderately abundant endoplasmic reticulum has a more peripheral distribution than in the promonocyte, and modest numbers of mitochondria (m) are present. Numerous pseudopodia (ps) extend from the cell surface. The peripheral lacunae (l) represent a tangential section through surface irregularities ×16,200. From Nicholls BA, Bainton DF: Differentiation of human monocytes in bone marrow and blood. Sequential formation of two granule populations. Lab Invest 1973;29:27-40, with permission. |
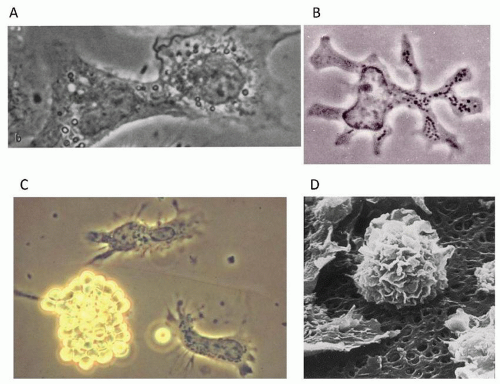 FIGURE 10.3. Morphology of mouse mononuclear phagocytes. A: Mouse peritoneal macrophages in culture on cover glasses undergo cell fusion shortly after addition of UV-irradiated Sendai virus. Note cytoplasmic bridge formation in nascent homokaryon. Phase contrast microscopy reveals characteristic nuclear morphology, rudimentary nucleoli, and extensive phase-lucent and phase-dense vesicles. Plasma membrane shows spreading of organelle-poor cell periphery with ruffling. From Gordon S, Cohn Z. Macrophage-melanocyte heterokaryons. I. Preparation and properties. J Exp Med 1970:131:981-1003, with permission. B,C: Phase contrast micrograph of dendritic cell isolated by Steinman and Cohn from mouse spleen. B: Note extensive dendrites, euchromatin nucleus, and mitochondria-rich cytoplasm, a feature of their high motility. C: Note lack of resetting of DCs with antibody-coated sheep erythrocytes, compared with macrophages in the same preparation, one of the features which allowed separation and their distinction. From Steinman RM, Cohn ZA. Identification of a novel cell type in peripheral lymphoid organs of mice. I. Morphology, quantitation, tissue distribution. J Exp Med 1973:1142-1162, with permission. D: Scanning electron microscopy of human alveolar macrophages cultured on cellulose membrane in a biphasic cell culture system. Note characteristic rounded appearance with extensive ruffles. From Wallaert B, Fahy O, Tsicopoulos A, Gosset P, Tonnel AB. Experimental systems for mechanistic studies of toxicant induced lung inflammation. Toxicol Lett 2000;112-113:157-163, with permission. |
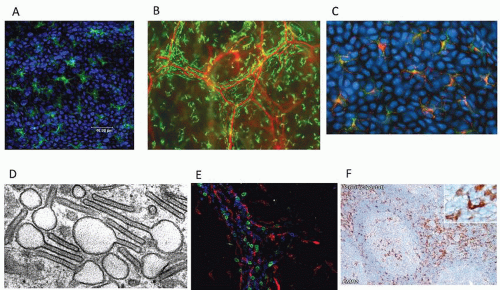 FIGURE 10.4. Mononuclear phagocytes in human skin and thymus. A: Epidermis: CD1a+ Langerhans cells. B: Dermis/interstitium Lyve-1+ macrophages and CD31+ endothelium. C: Epidermis: Langerhans cells express langerin and MHCII, which co-localize. D: Electron micrograph of Birbeck granules. E: Dermis: Triple labeling shows CD11c (DCs, green), XIIIa (macrophages, red) and CD3 (T cells, blue). F: Thymic macrophages and DCs express EMR2. Photomicrograph D from Romani N, Clausen BE, Stoitzner P. Langerhans cells and more: langerin expressing cell subsets in the skin. Immunol Rev 2010;234(1):120-141, with permission. Other images from M. Collin, N. Romani and T. Marafioti. |
TABLE 10.1 SELECTED PROPERTIES OF HUMAN MONONUCLEAR PHAGOCYTES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
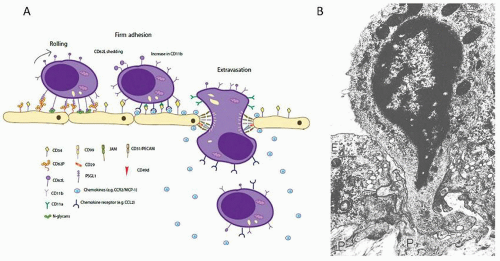 FIGURE 10.5. (A) Recruitment. Stages of monocyte adherence to endothelium and diapedesis, induced by inflammatory stimuli. The model is mainly based on the recruitment of neutrophils, with which it shares many features, although monocyte- specific chemokines, receptors, and adhesion ligands exist, especially in constitutive and noninfectious, metabolic forms of inflammation. Figure provided by S. Yona. (B) Monocyte diapedesis. Electron micrograph from collection of H. Florey. |
the pathophysiology of many human diseases including atherogenesis and other inflammatory diseases. The origin of steady state tissue macrophages and DCs is much less certain, as discussed below. It is likely that blood monocytes and blood DCs contribute, but circulating hematopoietic progenitor cells may also give rise to tissue leukocytes.
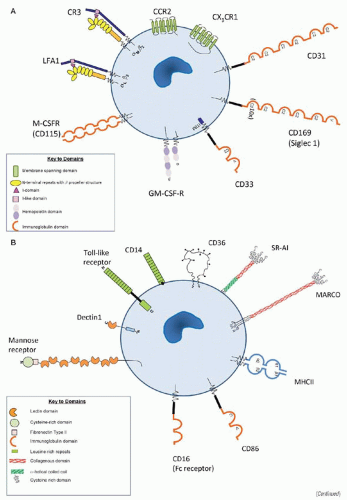 FIGURE 10.6. Selected macrophage plasma membrane receptors. (A) Growth, differentiation, adhesion and migration. (B) Recognition and sensing. |
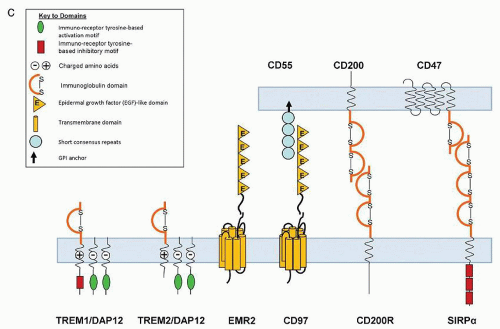 FIGURE 10.6. Selected macrophage plasma membrane receptors. (C) Potentiation and negative regulation. |
TABLE 10.2 SELECTED ANTIGEN MARKERS EXPRESSED BY HUMAN MONONUCLEAR PHAGOCYTES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
TABLE 10.3 SELECTED PLASMA MEMBRANE PATTERN RECOGNITION RECEPTORS (PRRs) EXPRESSED BY HUMAN MONONUCLEAR PHAGOCYTES | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
a related SR, and CD36, another promiscuous adhesion molecule also present on platelets and endothelial cells. The lectins include Dectin-1, the receptor for fungal β-glucan, expressed by monocytes, macrophages, neutrophils, and DCs; the mannose receptor, a multilectin with distinct carbohydrate recognition domains for GlcNac, mannosyl, and fucosyl ligands on microbes and selected host molecules; and a cysteine-rich domain which recognizes endogenous sulfated sugars. Other mannose binding lectins include DC-SIGN and Dectin-2; galectin is a receptor for galactosyl recognition. Additional plasma membrane receptors include CD163, an SR-like molecule that recognizes haptoglobin – hemoglobin complexes, and which is induced by glucocorticoids. Additional SR, not shown, are present on endothelial cells, which also express SR-A. Apart from the scavenger receptors, several other receptors can recognize apoptotic cells, e.g., the Tyro3, Axl, and Mer (TAM) receptors, which recognize Gas 6 and Protein S ligands. Several TIM molecules and CD300A bind phosphatidyl serine, another ligand for apoptotic cell recognition.
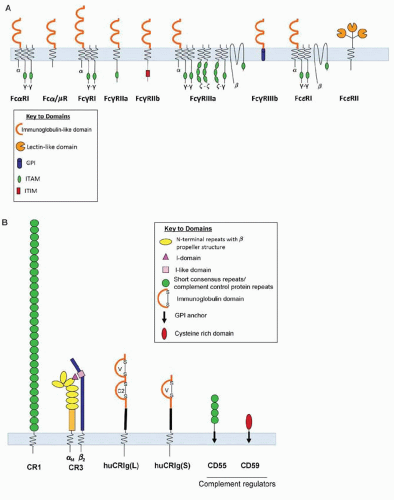 FIGURE 10.7. A: Fc receptors of human macrophages show diversity for Ig class ligands and signaling. See references for further details. B: Macrophage receptors for complement recognition and regulation. |
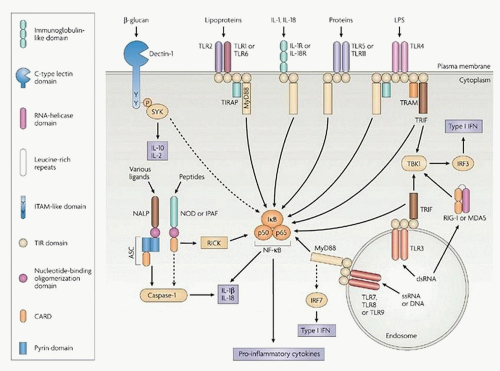 FIGURE 10.8. Cooperation of Toll-like receptor signals and Dectin-1 (with cytosolic inflammasome activation and type I interferon) in innate immune defense. From Trinichieri G, Sher A. Cooperation of Toll-like receptor signals in innate immune defense. Nat Immunol Rev 2007;7:179-190, with permission. |
disorders, and their dramatic amelioration by IL-1 antagonists. Inflammasomes are protein complex platforms which are required for the activation of inflammatory caspases and the maturation of their pro-inflammatory cytokines including IL-1β and IL-18. They are constructed around several proteins, including NLRP3, NLRC4, AIM2, and NLRP6, and recognize inflammatory signals arising from receptors for PAMPs, damage-associated molecules (DAMPs), and sterile particulates, such as uric acid crystals. Recognition of immune signals by one of several families of receptors results in direct activation of caspase-1 and caspase-5, secretion of potent pro-inflammatory cytokines, and a form of cell death called pyroptosis. PRRs of innate immune cells can be classified into phagocytic and sensor PRRs and, in addition to the plasma membrane receptors described above, include intracellular TLRs, retinoic acid-inducible gene I-like receptors (RLRs), and nucleotide-binding oligomerization domain-like receptors (NLRs). Inflammasome-mediated processes are important during microbial infections and also in regulating both metabolic processes and mucosal immune responses. For example, the NLRP3 inflammasome has
been demonstrated to be involved in antibacterial, viral, fungal, and parasitic immune responses. On infection with influenza A, endosomal TLR7 recognizes viral RNA and induces transcription of the NLRP3 inflammasome components. Selected bacteria have been shown to allow the cytoplasmic entry of flagellin, the NLRC4 ligand, leading to activation of the NLRC4 pathway. Some inflammasome activators are exogenous particles, e.g., silica and asbestos, whose uptake by pulmonary macrophages activates NLRP3 inflammasome-dependent caspase activation, cytokine, and cellular reactive oxygen species release, contributing to silicosis or asbestosis. Inflammasomes have also been implicated in metabolic pathologies with activation of caspase-1 by NLRP3 in adipose tissue, resulting in inhibition of insulin signaling, expression of TNF-α, and induction of CD4 T helper cells. However, a protective role for activated inflammasomes in age-related macular degeneration has also been proposed, since lack of NLRP3 or IL-18 exacerbates choroidal neovascularization.
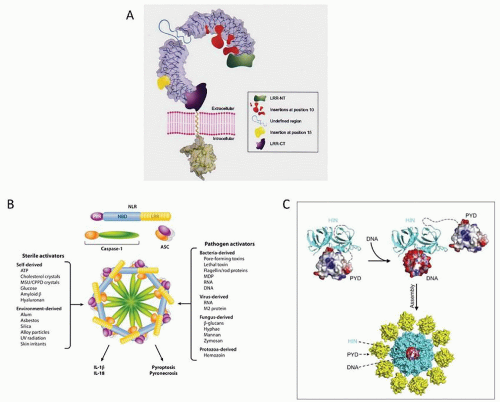 FIGURE 10.9. Models of inflammasome structure. A: A Toll-like receptor (TLR). TLRs are integral membrane glycoproteins with an N-terminal ectodomain and a single transmembrane domain. The ectodomain of a TLR7, TLR8, and TLR9 family member is depicted, with the leucine-rich repeat (LRR) solenoid shown with a gray molecular surface, and the N- and C-terminal flanking regions shown in green and purple, respectively. An undefined region present in TLR7, TLR8, and TLR9, but not in the other TLRs, is shown as a light blue string. Insertions within LRRs at position 10 are indicated in red and might contribute to the formation of the pathogen-associated molecular pattern (PAMP) binding site. An insert at position 15 is indicated in yellow. Also shown is a cartoon of the transmembrane domain (presumed to be a single α-helix), followed by a molecular surface representation of the TLR1 Toll-IL-1 and IL-18 receptor (TIR) domain. From Bell JY, Mullen GE, Leifer CA, Mazzoni A, Davies DR, Segal DM. Leucine-rich repeats and pathogen recognition in Toll-like receptors. Trends Immunol 2003;24:528-533, with permission. B: Activators of the inflammasome. Sterile activators include host- and environment-derived molecules, and pathogen-associated activators include PAMPs derived from bacteria, viruses, fungi, and protozoa. Assembly of the NLRs, ASC, and caspase-1 leads to the formation of a pentad- or heptamer structure: the inflammasome. Activation of the inflammasome leads to maturation and secretion of IL-1β and IL-18, as well as inflammatory cell death, by either pyroptosis or pyronecrosis. ASC, apoptosis-associated speck-like protein containing a caspase recruitment domain; CPPD, calcium pyrophosphate dihydrate; MDP, muramyl dipeptide; MSU, monosodium urate; NLR, nucleotide-binding domain, leucine-rich repeat containing; PAMP, pathogen-associated molecular patterns. From Davis BK, Wen H, Ting JP. The inflammasome NLRs in immunity, inflammation, and associated diseases. Annu Rev Immunol 2011;29:707-735. C: Electrostatic attraction underlies innate dsDNA recognition by the HIN domains. Both oligonucleotide/oligosaccharide binding folds and the linker between them engage the dsDNA backbone. An autoinhibited state of AIM2 is activated by DNA that liberates the PYD domain. DNA serves as an oligomerization platform for the inflammasome assembly. From Jin T, Perry A, Jiang J, et al. Structures of the HIN domain: DNA complexes reveal ligand binding and activation mechanisms of the AIM2 inflammasome and IFI16 receptor. Immunity 2012;36(4):561-571, with permission. |
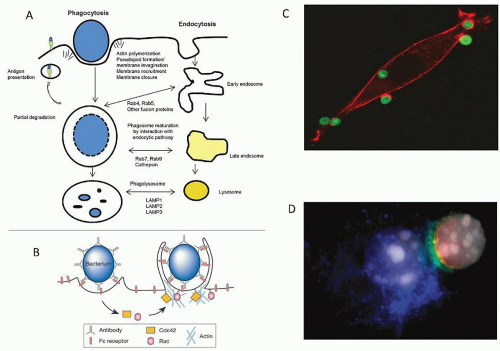 FIGURE 10.10. A: Phagocytosis and endocytosis pathways. Particulates are taken up by actin-dependent sequential maturation processes, involving membrane fusion and fission, which intersect with the endocytic pathway at several stages. Cytosolic small guanosine triphosphatases (rabs) determine organelle-specific interactions. Membrane is recycled to the plasma membrane, with processed antigen. Progressive acidification and delivery of lysosomal hydrolases result in terminal degradation. Compartment membranes express marker proteins such as LAMP1; the pan-macrophage CD68 antigen is associated with late endosomes and lysosomes. B: Schematic representation of FcR-mediated uptake of a bacterium by a zipper-like mechanism, initiating actin assembly. Modified from Conner SD, Schmid SL. Regulated portals of entry into the cell. Nature 2003;422(6927):37-44. C: Dectin-1 transfected fibroblast taking up unopsonized zymosan (yeast wall particle) through β-glucan recognition, to initiate a phagocytic cup. (Courtesy of G. Brown.) D: Uptake of Pneumocystis carinii via Dectin-1. Contact zone resembles phagocytic synapse described by Goodridge et al.30 |
by Dectin-1, which has a hemi-ITAM in its cytoplasmic domain, by a syk- and CARD9-dependent pathway. Uptake is associated with acidification, to promote digestion of all macromolecules.
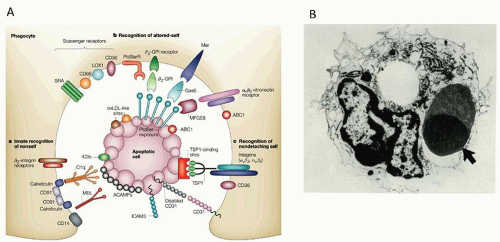 FIGURE 10.11. A: Phagocytic receptors for apoptotic cell phagocytosis. Macrophages and immature myeloid dendritic cells (mDCs) express multiple receptors for direct or opsonic binding, e.g., mannose binding lectin (MBL) or milk fat globulin (mFGE8). Phosphatidylserine (PS) exposed on the outer surface of the apoptotic cell can be recognized by a range of receptors, including TIM4, TIM1 and CD300A (not shown). Discrimination of nonself and altered-self may involve combinations of different phagocyte receptors. Apoptotic cell uptake results in an antiinflammatory response (e.g., release of transforming growth factor-β and prostaglandin E2), but has also been implicated in cross-presentation by DCs. From Savill J, Dransfield I, Gregory C, Haslett C. A blast from the past: clearance of apoptotic cells regulates immune responses. Nat Rev Immunol 2002;2:965-975, with permission. B: Thymic macrophage with apoptotic thymocyte in phagocytic vacuole. The vast majority of thymocytes never leave the thymus, undergo apoptosis, and are rapidly removed by macrophages (×4,500). |
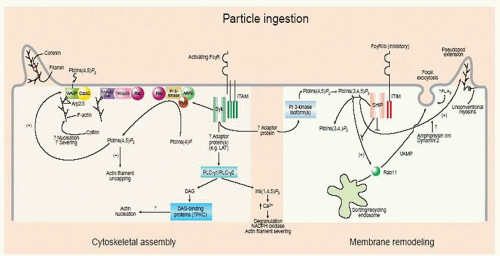 FIGURE 10.12. Plasma membrane remodeling, cytoskeletal assembly, and cytoplasmic signaling associated with Fc receptor-mediated particle ingestion. From Greenberg S. Diversity in phagocytic signalling. J Cell Sci 2001;114(Pt 6):1039-1040, with permission.44 |
DCs are characterized by high expression of CD141 in humans and CD8 in mice. They also possess a unique chemokine receptor XCR1 and lectin CLEC9A (DNGR-1) with actin as ligand; and they express a high level of TLR3. Specialized biochemical machinery allows these cells to transfer exogenous antigen to the endogenously loaded MHC class I presentation pathway.
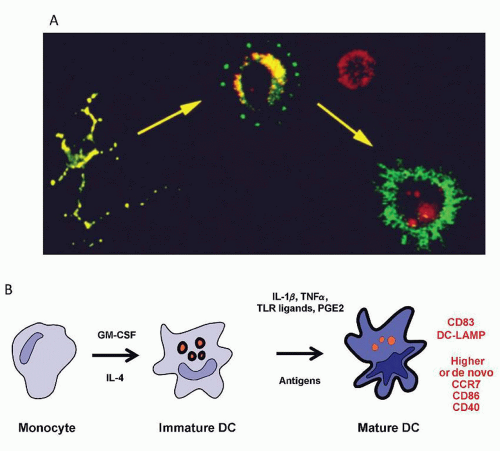 FIGURE 10.13. Maturation/activation of dendritic cells (DCs) leads to acquisition of APC function. A: DC maturation ex vivo. Langerhans cells upon explantation from mouse skin undergo “spontaneous” maturation with redistribution of MHCII from an intracellular compartment to the cell surface. (Courtesy of Ira Mellman and Ralph Steinman.) B: One of the first systems to produce large numbers of human dendritic cells from blood monocytes; the added cytokines and other maturation stimuli influence the function of the DCs. Based on studies by Lanzavecchia, Sallusto, Romani, Schuler, Bhardwaj, and their colleagues.
Related posts:Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
 Get Clinical Tree app for offline access
Get Clinical Tree app for offline access

|