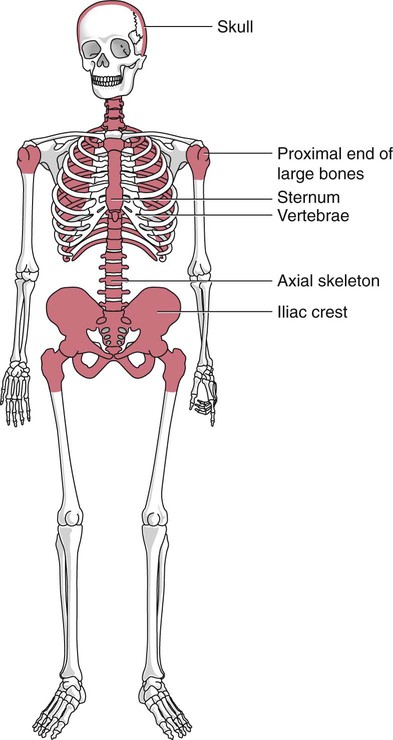
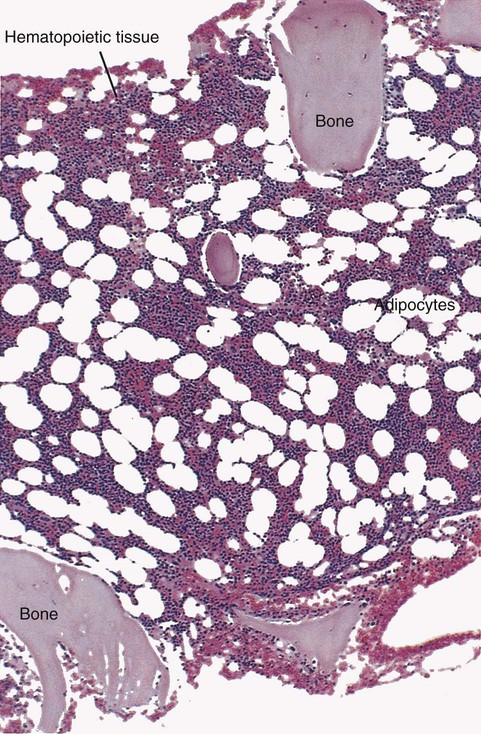
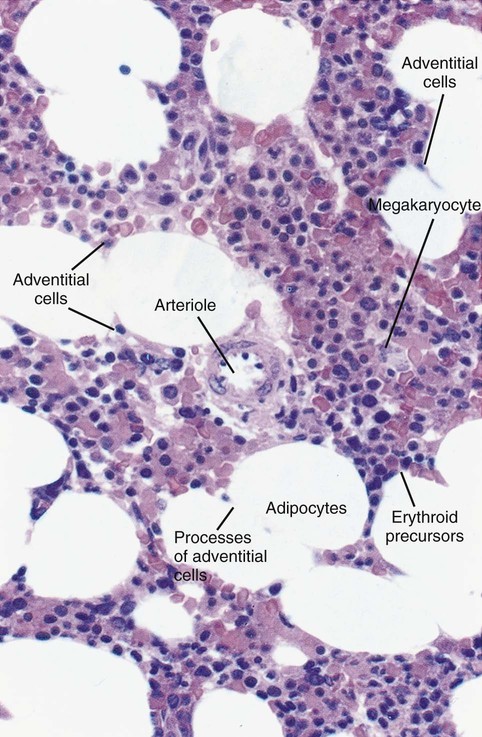
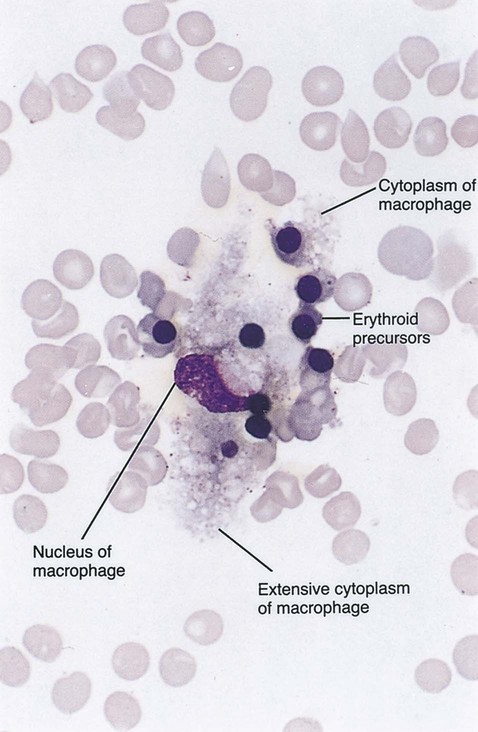
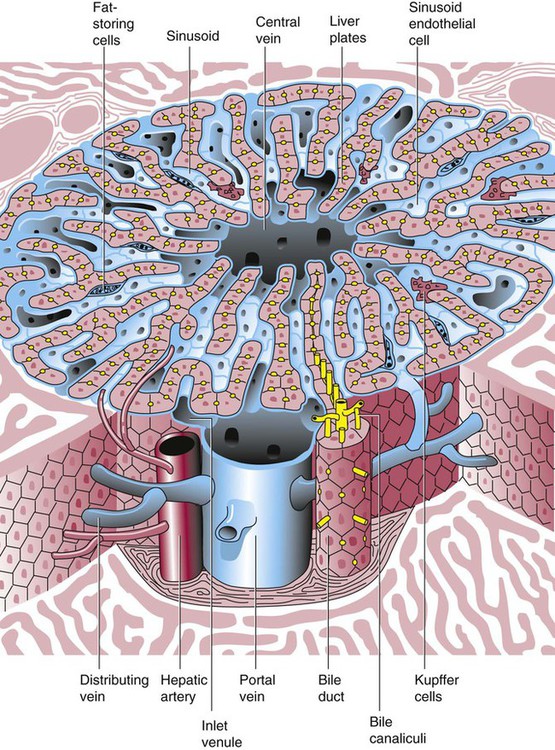
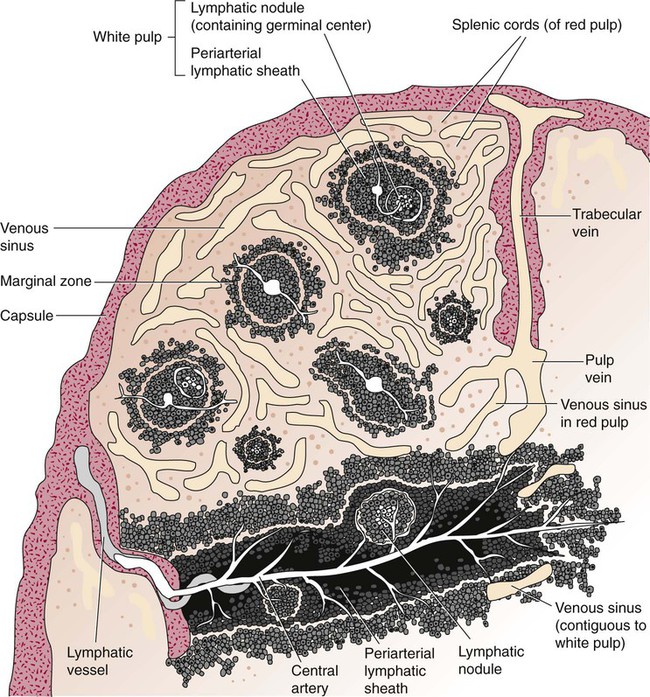
After completion of this chapter, the reader will be able to: 2. Discuss the evolution and formation of blood cells from embryo to fetus to adult, including anatomic sites and cells produced. 3. Predict the likelihood of encountering active marrow from biopsy sites when given the patient’s age. 4. Relate normal and abnormal hematopoiesis to the various organs involved in the hematopoietic process. 5. Explain the stem cell theory of hematopoiesis, including the names of various stem cells and progenitor cells and their lineage associations. 6. Discuss the roles of hematopoietic growth factors in differentiation and maturation sequences of hematopoietic progenitor cells, including nonspecific and lineage-specific factors. 7. Describe general morphologic changes that occur during cell maturation. 8. Define apoptosis and discuss the relationship between apoptosis, growth factors, and stem cell differentiation. 9. Discuss the roles of various cytokines and hematopoietic growth factors in the process of hematopoiesis. 10. Discuss therapeutic applications of cytokines and hematopoietic growth factors. Hematopoiesis is considered to begin around the nineteenth day of embryologic development after fertilization. Progenitor cells of mesenchymal origin migrate from the aorta-gonad-mesonephros region of the developing aorta-splanchnopleure to the yolk sac.1 The cells arising from the aorta-gonad-mesonephros region give rise to hematopoietic stem cells (HSCs), but not to primitive erythroblasts. The primitive erythroblasts found in the yolk sac arise from mesodermal cells, which initially line the cavity of the yolk sac. These primitive cells migrate from the periphery into the central cavity of the yolk sac, where they develop into primitive erythroblasts.2–5 The remaining cells surrounding the cavity of the yolk sac are called angioblasts and form the future blood vessels.2–5 The yolk sac phase of hematopoiesis is characterized by the development of primitive erythroblasts that produce measurable amounts of hemoglobins, including Portland, Gower-1, and Gower-2 (see Chapter 10). Yolk sac hematopoiesis does not contribute significantly to definitive hematopoiesis.4 This phase of hematopoiesis occurs intravascularly, or within a developing blood vessel. The hepatic phase of hematopoiesis begins at 4 to 5 gestational weeks and is characterized by recognizable clusters of developing erythroblasts, granulocytes, and monocytes.5 The developing erythroblasts signal the beginning of definitive hematopoiesis with a decline in primitive hematopoiesis of the yolk sac. In addition, lymphoid cells begin to appear.6,7 Hematopoiesis during this phase occurs extravascularly, with the liver remaining the major site of hematopoiesis during fetal life and retaining activity until 1 to 2 weeks after birth. Hematopoiesis in the aorta-gonad-mesonephros region and the yolk sac disappears during this stage. Hematopoiesis in the fetal liver reaches its peak by the third month of development (Figure 7-1). The developing spleen, kidney, thymus, and lymph nodes contribute to the hematopoietic process during this phase. The thymus, the first fully developed organ in the fetus, becomes the major site of T-cell production, whereas the kidney and spleen produce B cells. Production of megakaryocytes also begins during the hepatic phase. The spleen gradually decreases granulocytic production and involves itself solely in lymphopoiesis. During the hepatic phase, detectable levels of hemoglobin (Hb) F, Hb A, and Hb A2 may be present.8 During the fifth month of fetal development, hematopoiesis begins in the developing bone marrow cavity. This transition is called medullary hematopoiesis because it occurs in the medulla or inner part of the bone marrow. During this phase, mesenchymal cells, which are a type of embryonic tissue, migrate into the core of the bone and differentiate into skeletal and hematopoietic blood cells.9,10 Hematopoietic activity, especially myeloid activity, is apparent during this stage of development, and the myeloid-to-erythroid ratio approaches the adult level of 3 : 1 by 21 weeks of gestation.11 By the end of the sixth month, the bone marrow becomes the primary site of hematopoiesis. Measurable levels of erythropoietin (EPO), granulocyte colony-stimulating factor (G-CSF), granulocyte-monocyte colony-stimulating factor (GM-CSF), fetal hemoglobin, Hb A2, and adult hemoglobin can be detected. In addition, cells at various stages of maturation can be seen in all three lineages. A central space is created within the cavities of these bones by resorption of cartilage and endosteal bone. Mesenchymal cells migrate into the space and eventually differentiate into three cell types, which give rise to the blood and bone marrow matrix cells (reticular cells and adipose tissue). Reticular cells are formed on the exterior surfaces of the venous sinuses and extend long, narrow branches into the perivascular space, creating a meshlike network; this provides a supportive skeletal network for developing hematopoietic cells, macrophages, and mast cells.10 During infancy and early childhood, the bone marrow consists primarily of red active marrow. Between 5 and 7 years of age, adipocytes become more abundant and begin to occupy the spaces in the long bones previously dominated by active marrow. The process of replacing the active marrow by adipose tissue (yellow marrow) during development is called retrogression and eventually results in restriction of the active marrow to the flat bones, sternum, vertebrae, pelvis, ribs, skull, and proximal portion of the long bones (Figure 7-2). Areas located within the bone marrow cavity where red marrow has been replaced by yellow marrow consist of an mixture of adipocytes, undifferentiated mesenchymal cells, and macrophages. Inactive yellow marrow is also scattered throughout active red marrow and is capable of reverting back to active marrow in cases of increased demand on the bone marrow.8 Such cases might be excessive blood loss or increased erythrocyte destruction in the bone marrow by toxic chemicals or irradiation. The red marrow is composed of extravascular cords that contain all of the developing blood cell lineages, stem and progenitor cells, adventitial cells, and macrophages (Figures 7-3 and 7-4). The cords are separated from the lumen of the sinusoids by endothelial and adventitial cells and are located between the trabeculae of spongy bone. Trabeculae are projections of calcified bone radiating out from the cortical bone into the marrow space and provide support for the developing marrow. The hematopoietic cells tend to develop in specific niches within the cords. Normoblasts develop in small clusters adjacent to the outer surfaces of the vascular sinuses (Figure 7-5); in addition, some normoblasts are found surrounding iron-laden macrophages (Figure 7-6). Megakaryocytes are located close to the vascular walls of the sinuses, which facilitates the release of platelets into the lumen of the sinusoids. Immature myeloid (granulocytic) cells through the metamyelocyte stage are located deep within the cords. As these maturing granulocytes proceed along their differentiation pathway, they move closer to the vascular sinuses.9 As blood cells come in contact with endothelial cells, they bind to the surface via a receptor-mediated process. Cells pass through pores in the endothelial cytoplasm and are released into the circulation.12 The nutrient and gas requirements of the marrow are supplied by the nutrient and periosteal arteries, which enter via the bone foramina. The nutrient artery supplies blood only to the marrow.10 It coils around the central longitudinal vein, which passes along the bone canal. In the marrow cavity, the nutrient artery divides into ascending and descending branches that also coil around the central longitudinal vein. The arteriole branches that enter the inner lining of the cortical bone (endosteum) form sinusoids (endosteal beds), which connect to periosteal capillaries that extend from the periosteal artery.13 The periosteal arteries provide nutrients for the osseous bone and the marrow. Their capillaries connect to the venous sinuses located in the endosteal bed, which empty into a larger collecting sinus that opens into the central longitudinal vein.14 Blood exits the marrow via the central longitudinal vein, which runs the length of the marrow. The central longitudinal vein exits the marrow through the same foramen where the nutrient artery enters. Hematopoietic cells located in the endosteal bed receive their nutrients from the nutrient artery. The hematopoietic inductive microenvironment plays an important role in stem cell differentiation and proliferation.15,16 It is responsible for supplying a semifluid matrix, which serves as an anchor for the developing hematopoietic cells. The matrix is responsible for maintaining differentiation and proliferation and provides a supporting tissue in the hematopoietic inductive microenvironment. Stromal cells in the matrix are of several types: (1) endothelial cells, (2) adipocytes, (3) macrophages, (4) osteoblasts, (5) osteoclasts, and (6) reticular cells (fibroblasts). Endothelial cells are broad flat cells that form a single continuous layer along the inner surface of the bone marrow sinus.15 They regulate the flow of particles entering and leaving hematopoietic spaces. Adipocytes are large cells with a single fat vacuole; they secrete various steroids that influence erythropoiesis and maintain bone integrity.14 They also play a role in regulating the volume of the marrow in which active hematopoiesis occurs.14 Macrophages function in phagocytosis and secretion of various cytokines that regulate hematopoiesis and are located throughout the marrow space.17 Other cells involved in cytokine production include endothelial cells, adipocytes, and fibroblasts. Osteoblasts are bone-forming cells, and osteoclasts are bone-resorbing cells. Reticular cells are associated with the formation of reticular fibers that form a lattice that supports the vascular sinuses and developing hematopoietic cells. Stromal cells are believed to be derived from fibroblasts. They play a role in support and regulation of hematopoietic stem/progenitor cell survival and differentiation.15 The extracellular matrix of the bone marrow contains proteoglycans or glycosaminoglycans, fibronectin, collagen, laminin, hemonectin, and thrombospondin.15 Proteoglycans are expressed on the endothelial cell surface and mediate progenitor binding to the stroma. Fibronectin, collagen, laminin, hemonectin, and thrombospondin function as adhesion molecules, promoting the adhesion of HSCs to the extracellular matrix. The liver consists of two lobes situated beneath the diaphragm in the abdominal cavity. The position of the liver with regard to the circulatory system is optimal for gathering, transferring, and eliminating substances via the bile.18 Anatomically, liver cells are arranged in radiating hepatic lobules emanating from a central vein (Figure 7-7). Adjacent to the longitudinal lobes of the liver and separated only by a small space are sinusoids, which are lined by two types of cells: Kupffer cells and epithelial cells. Kupffer cells are macrophages, removing cellular and foreign debris from the blood that circulates through the liver; they also are responsible for protein synthesis.19 The epithelial cells are arranged in the lining so as to be separated from one another by a noncellular area; this arrangement allows plasma to have direct access to the hepatocytes. This unusual organization of the liver and its location in the body enables it to be involved in many varied functions. The liver is often involved in blood-related diseases. In porphyrias, the liver exhibits enzymatic deficiencies that result in the accumulation of the various intermediary porphyrins. In severe hemolytic anemias and red blood cell (RBC) dysplasias, the conjugation of bilirubin and the storage of iron are increased. The liver sequesters membrane-damaged RBCs and removes them from the circulation. The liver is capable of extramedullary hematopoietic production in case of bone marrow shutdown.14 It is directly affected by storage diseases of the monocyte/macrophage (Kupffer) cells as a result of enzymatic deficiencies that cause hepatomegaly with ultimate dysfunction of the liver (Gaucher disease, Niemann-Pick disease, Tay-Sachs disease; see Chapter 28). The spleen is the largest lymphoid organ in the body. The spleen is located directly beneath the diaphragm behind the fundus of the stomach in the upper left quadrant of the abdomen. It is vital but not essential for life and functions as an indiscriminate filter of the circulating blood. In a healthy individual, the spleen contains about 350 mL of blood.18 The exterior surface of the spleen is surrounded by a layer of peritoneum and inwardly by a connective tissue capsule. The capsule projects inwardly, forming trabeculae that divide the spleen into discrete regions. Located within these regions are three types of splenic tissue: (1) white pulp, (2) red pulp, and (3) a marginal zone. The white pulp consists of scattered follicles with germinal centers containing lymphocytes, macrophages, and dendritic cells. Aggregates of lymphocytes surround splenic arteries that pass through these germinal centers. Adjacent to the splenic arteries is a region called the periarteriolar lymphatic sheath. This area consists of lymphoid nodules containing primarily B lymphocytes. Activated B lymphocytes are found in the germinal centers.19 The marginal zone surrounds the white pulp and forms a reticular meshwork containing blood vessels, macrophages, and specialized B cells. The red pulp is composed primarily of vascular sinusoids and sinuses separated by cords of tissue (cords of Billroth) containing specialized macrophages that are loosely connected to the dendritic process, creating a spongelike region that functions as a filter for blood passing through the region.19 As RBCs pass through the cords of Billroth, there is a decrease in the flow of blood, which leads to stagnation and depletion of the RBCs’ glucose supply. These cells are subject to increased damage and stress that may lead to their removal from the spleen. The spleen uses two methods for removing senescent RBCs from the circulation: (1) culling, in which the cells are phagocytosed with subsequent degradation of cell organelles, and (2) pitting, in which splenic macrophages remove inclusions or damaged surface membrane from the circulating RBCs. The spleen synthesizes immunoglobulin M in the germinal centers, and it serves as a storage site for platelets. In a healthy individual, approximately 30% of the total platelet count is sequestered in the spleen.20 The spleen has a rich blood supply, receiving approximately 350 mL/min. Blood enters the spleen through the central splenic artery located at the hilum and branches outward through the trabeculae. The branches enter all three regions of the spleen: the white pulp with its dense accumulation of lymphocytes, the marginal zone, and the red pulp. The venous sinuses, which are located in the red pulp, unite and leave the spleen as splenic veins (Figures 7-8 and 7-9).21 In the rapid-transit pathway, blood cells enter the splenic artery and pass directly to the sinuses in the red pulp and continue to the venous system to exit the spleen. When splenomegaly occurs, the spleen becomes enlarged and is palpable. This occurs as a result of many conditions, such as chronic leukemias, genetic defects in RBCs, hemoglobinopathies, Hodgkin disease, thalassemia, malaria, and the myeloproliferative disorders. Often splenectomy is beneficial in cases of excessive destruction of RBCs, such as severe hereditary spherocytosis, storage disorders, and autoimmune hemolytic anemias, when treatment with corticosteroids does not effectively suppress hemolysis.22,23 Splenectomy also may be indicated in severe cases of agnogenic myeloid metaplasia associated with splenomegaly, severe refractory hemolytic anemia, thrombocytopenia, or qualitative platelet function defect syndromes.22,23 After splenectomy, platelet and leukocyte counts increase transiently.22 In sickle cell anemia, repeated splenic infarcts caused by sickled RBCs trapped in the small-vessel circulation of the spleen cause tissue damage and necrosis, which often results in autosplenectomy (see Chapter 26). Hypersplenism is an enlargement of the spleen resulting in some degree of pancytopenia despite the presence of a hyperactive bone marrow. The most common cause is congestive splenomegaly secondary to cirrhosis of the liver and portal hypertension. Other causes include thrombosis, vascular stenosis, other vascular deformities such as aneurysm of the splenic artery, and cysts.24 Lymph nodes are organs of the lymphatic system located along the lymphatic capillaries that parallel, but are not part of, the circulatory system. The nodes are bean-shaped structures (1 to 5 mm in diameter) that occur in groups or chains at various intervals along lymphatic vessels. They may be superficial (inguinal, axillary, cervical, supratrochlear) or deep (mesenteric, retroperitoneal). Lymph is the fluid portion of blood that escapes into the connective tissue and is characterized by a low protein concentration and the absence of RBCs. Afferent lymphatic vessels carry circulating lymph to the lymph nodes. Lymph is filtered by the lymph nodes and exits via the efferent lymphatic vessels located in the hilus of the lymph node.25 Similar in structure to the spleen, lymph nodes consist of an outer capsule that forms trabeculae and provides support for macrophages and the predominant population of lymphocytes. Lymph nodes can be divided into two basic regions: an outer region called the cortex and an inner region called the medulla. Trabeculae radiate through the cortex and the medulla, dividing the interior of the lymph node into specific areas (Figure 7-10). In the cortical region, these areas are known as cortical nodules and contain follicles. These follicles contain foci of B-cell proliferation termed germinal centers.9
Hematopoiesis
Hematopoietic Development
Mesoblastic Phase (Yolk Sac Phase)
Hepatic Phase
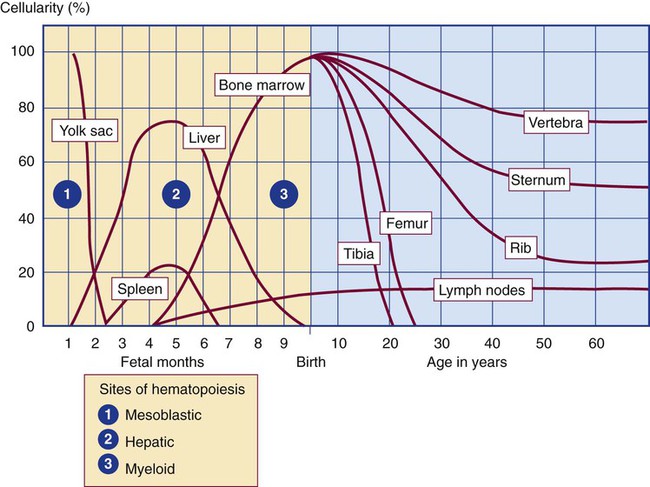
Medullary (Myeloid) Phase
Adult Hematopoietic Tissue
Bone Marrow
Red Marrow
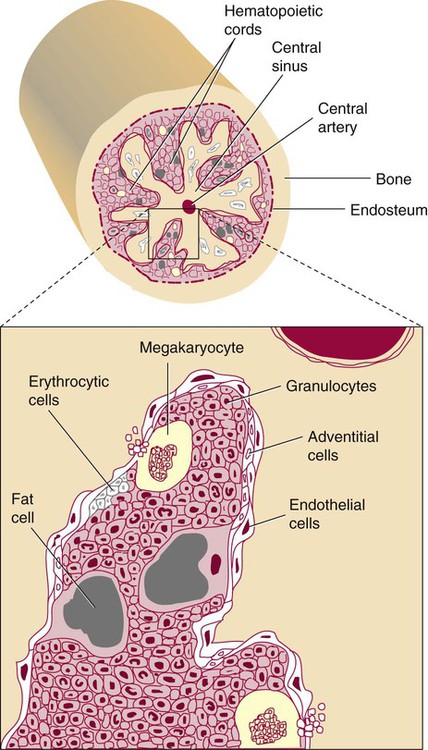
Marrow Circulation
Hematopoietic Microenvironment
Liver
Liver Pathophysiology
Spleen
Spleen Pathophysiology
Lymph Nodes
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Hematopoiesis