Fig. 23.1
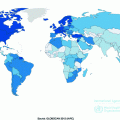
a A comparison of age-standardized mortality rates (per 100,000) of HCC among men, across different countries. Data are taken from the Globocan 2012 database. b A comparison of age-standardized mortality rates (per 100,000) of HCC among women, across different countries. Data are taken from the Globocan 2012 database
23.2.1 Trends by Geography
The age-standardized incidence rates of HCC show tremendous geographic differences, with a relatively low incidence in Europe and North America (2–7 per 100,000 cases per year), and a higher incidence in East Asia and Africa (>20 per 100,000 cases per year) (Fig. 23.1a, b) [1]. More than half (N = 395,000) of the cases of HCC occur in China, where it is the second most common cancer, primarily due to the endemic presence of the Hepatitis B Virus (HBV) (described under risk factors below) [4]. In Japan, the high incidence is due to the high prevalence of Hepatitis C virus (HCV) among the population [5].
More than 80 % of all cases occur in developing countries. In Africa HCC is the 4th most common cancer, with a particularly high incidence in the sub-Saharan area [4, 5]. The age of onset of HCC also varies considerably. In North America, the mean age at the diagnosis of HCC is 60 years, in Africa 35 years, and in Taiwan around 50 years [1]. In low-risk populations, the highest incidence of HCC is generally recorded among individuals of 75 years or older. HCC is rarely seen during the first 4 decades of life, except in population where infection with HBV is endemic.
23.2.2 Prognosis and Trends in Mortality
HCC is a highly lethal cancer, and thus the mortality rates of HCC shows the same geographic distribution of incidence [6]: over the last decades HCC mortality has decreased in Southern European countries, but increased in former low mortality areas of central and Northern Europe. Trends in HCC mortality in the United States (U.S.) and Australia were similar to those observed in central and Northern Europe, with an increased mortality rate in individuals 45–64 years. These mortality changes across Europe could be related to the decrease of HBV infection rates after the adoption of vaccination programs and the reduction of alcohol intake and tobacco smoking in Southern Europe. On the other hand, the increasing prevalence of HCV infection and alcohol consumption explains the opposite trend in central and Northern Europe. The increase of diabetes and the consequent overweight and obesity in several populations may have had a role in the recent adverse outcomes in HCC particularly in North America [7].
23.3 Pathogenesis and Risk Factors
HCC is a multistep process involving various risk factors, leading to chronic liver damage and genetic alterations. The most important single risk factor for HCC is cirrhosis, which can occur as a consequence of HBV and HCV infections as well as hemochromatosis and α1-antitrypsin deficit showed an independent risk of developing HCC [1]. Less studied independent risk factors include the exposure to aflatoxin and hormones, and recently also tobacco smoking has been shown to correlate with HCC [8].
23.3.1 Cirrhosis
Cirrhosis is a major health problem worldwide, and apart from the clinical problem of the end-stage disease, 80–90 % of HCC both in Asia and in Western Countries are estimated to be associated with this condition [9, 10]. The risk of HCC among individuals with cirrhosis ranges from 30 % at 5 years in Asian patients with HCV-driven cirrhosis, 20 % in patients with hemochromatosis, to 10 % for cirrhosis associated with alcohol and primary biliary disease [10]. In Western countries, alcohol consumption is a leading cause of cirrhosis, and its association with HCC is reinforced by other causes of chronic liver damage [11]. The oncogenic risk of cirrhosis largely derives from the hyperproliferative status of the regenerative hepatocytic nodules, but other molecular mechanisms have been proposed. For example, in cirrhotic liver tissue the activated stellate cells are able to activate both mitogen-activated protein kinase and the phoshatidylinositol-3 kinase pathways in response to circulating and paracrine pro-inflammatory cytokines (platelet-derived growth factor and transforming growth factor-ß, among others). Notably, the protein kinase and the phoshatidylinositol-3 kinase pathways represent the signal cascade typically seen in hepatocarcinogenesis [12].
In Western countries, with lower prevalence of HBV infection, 15–20 % of HCC arises without cirrhosis [13, 14]. HCC without cirrhosis is more likely occurring among men, has a bimodal distribution of age (peaking at 2nd and 7th decades) and a diagnosis typically at a symptomatic advanced stage, compared to patients with cirrhosis. Clinically, these HCC differ as well, including presenting as single large masses and lower serum α-fetoprotein level [13, 15]. Genomically, these cancers have a lower rate of p53 mutation and a higher prevalence of β[beta]-catenin mutation, p14 inactivation and global gene methylation [13].
Despite the higher stage at the diagnosis, patients with non-cirrhotic HCC have a better overall survival and disease-free survival than cirrhotic patients, due to the underlying liver conditions [14].
23.3.2 HBV
HBV is a Hepadnavirus infecting 350,000,000 people worldwide [5], representing the primary cause of liver cirrhosis and HCC in Asia, Africa, and South America, although incidence varies considerably across countries [4, 16]. In endemic areas such as China, HBV infection causes approximately 80 % of HCC cases [6]. Moreover, HBV is associated with virtually all HCC cases in children. In Western Countries where the mass vaccination against HBV has taken place since the 1980s, the incidence of HBV-related hepatitis and HCC has lowered [17].
Eight different HBV genotypes have been identified (A–H), with distinct geographical and ethnic distribution. In Asia end-stage liver disease, cirrhosis, and HCC are much more associated with the genotype C than genotype B; in Western Europe and North America genotype D has a greater incidence in severe liver disease and HCC than genotype A. Furthermore, recent data associated genotype B with an earlier onset of HCC and with a non-cirrhotic background [6]. The lifetime risk of developing HCC in patients with chronic HBV liver disease is 10–25 % [18], with a 20–30 years lag time from HBV infection to diagnosis of HCC [19].
Most HBV-positive patients develop HCC in a cirrhotic background, although there is evidence of a direct carcinogenic effect of HBV in non-cirrhotic patients: 20 % of all HBV-related HCC developed in liver without cirrhosis and, in some cases, even without chronic hepatitis [20]. Among the proposed mechanisms of this direct viral carcinogenicity are the integration of the double-stranded DNA of HBV in the genome of the infected hepatocytes and the binding of the product of the viral HBx gene to product of the p53 gene causing its inactivation [21, 22]. HBx does not bind the DNA but makes protein–protein interaction, triggering the transcriptional activation of several viral and cellular promoters and enhancers. HBx also deregulates the expression of oncogenes (e.g., c-Myc and c-Jun), cytokines and transcription factors, and modulates cytoplasmic signal transduction pathways, and is involved in a number of oncogenic effects [20, 21, 23–25]. It was recently shown that HBX may also have an important role in modulating the epigenetic control of viral and cellular genes, including a number of tumor suppressor genes [18].
HBV-related HCC commonly exhibits a higher rate of chromosomal abnormalities than liver tumors linked to other risk factor, and it has been suggested that HBV might generate genomic instability either through viral DNA integration or the activity of its proteins [18].
23.3.3 HCV
Hepatitis C virus (HCV) is a single-stranded RNA virus of the Flaviviridae family, with at least six known genotypes characterized by high genetic variability. The highest prevalence of HCV infection worldwide is recorded in the Middle East and Africa, the lowest in Europe and America [26]. In North America, Japan, and some European countries like Italy and Spain, HCV is the main risk factor of cirrhosis and HCC [18]. The incidence of HCC in patients infected with HCV was estimated up to 4 %, with a mean time from HCV infection to the diagnosis of HCC of 30 years [10, 27].
HCV-associated HCC arises almost invariably in livers with cirrhosis, since there do not appear to be direct carcinogenic roles in HCC [5] and the pathogenesis is more likely to be related to the HCV-driven chronic hepatitis and fibrosis. HCV leads to chronic inflammation, immune-mediate hepatocytes death, tissue damage, fibrosis activation by hepatic stellate cells, and replicative senescence due to telomerase shortening. In addition, factors such as oxidative stress, steatosis, and insulin resistance accelerate the evolution to cirrhosis and HCC [28] There is no evidence that the HCV integrates into the host genome, although HCV proteins interact with many host-cell proteins at a cytoplasmic level, thereby influencing cell signaling, transcription, proliferation, apoptosis, and translational regulation [29]. In vitro studies show a possible oncogenic role for four HCV proteins: core protein, NS3, NS5b, and NS5a [21, 23, 29, 30]. In particular, HCV core protein and NS3 are able to modulate the expression of p21 and p53, with alterations in the apoptosis and cell proliferation [23]. Another proposed mechanism for HCV carcinogenesis is an increased cellular oxidative stress by compromising the endoplasmic reticulum [23].
23.3.4 Hemochromatosis
Hemochromatosis is a relatively frequent metabolic disorder leading to iron accumulation and cirrhosis, and it is associated with an increased risk of developing HCC, compared to the general population [2]. HCC accounts for as many as 45 % of disease-related deaths in patients afflicted by hemochromatosis; the estimated risk of HCC in hemochromatosis is 8–10 % in most studies, although lower risk (as low as 1.7 %) has been recorded [31]. Although the effect of hemochromatosis is largely attributed to its effect on cirrhosis, some studies have found that intracellular iron overload can have a role in HCC pathogenesis [32]. HCC in hemochromatosis is commonly attributed to the iron overload in the hepatocytes that leads to direct oxidative damage by the creation of free radicals: this damage associated with an activation of stellate cells leads to liver fibrosis, cirrhosis, and then HCC [33]. Very recently, a “two-hit” hypothesis has been proposed from the observation of iron-free dysplastic nodules and HCC: according to this hypothesis the tumor initiation might be represented by the iron overload leading to cirrhosis and a second step, tumor promotion, might be represented by the proliferation of iron-free malignant hepatocytes in the effort to avoid the citotoxic effect of iron [28].
23.3.5 Aflatoxin B1
Aflatoxin B1 is a toxin produced by fungi of the Aspergillus family, and can be present in contaminated foods including wheat, nuts, and corn. It is found in moist climates, including parts of Asia and sub-Saharan Africa. Among the four known Aflatoxins (B1, B2, G1 and G2), Aflatoxin B1 is the most potent hepatocarcinogen in animal models [7].
Related posts:
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


