In the last two decades, several vector-borne infections have emerged as major causes of human morbidity and mortality. A dramatic recent example is the spread of West Nile virus (WNV) into the Western Hemisphere from endemic foci in the Middle East, Africa, and eastern Europe. Moreover, the recent epidemic activity of WNV has also increased and expanded, even in the areas where the virus previously was endemic.
Other examples of the spread of vector-borne infections include the emergence of all four serotypes of dengue virus in the Caribbean and Central and South America, the expansion of the geographic foci of Japanese encephalitis virus to western India and Pakistan, and the emergence of yellow fever in West Africa.1, 2 and 3 In addition, the incidence of Lyme disease in the United States and that of tick-borne encephalitis virus in eastern and central Europe and Russia have increased in the last decade.
Malaria remains a major cause of infectious morbidity and mortality in sub-Saharan Africa and other tropical countries in Asia, the Middle East, and South America. However, despite the continued major public health impact of malaria on morbidity and mortality, especially in infants and children, there appears to have been a reduction in the incidence of the disease in some areas in Africa in the past few years.
Although the emergence of each of these vectorborne infections is linked to unique changes in human activity and the environment, several cross-cutting factors facilitate the emergence of infectious diseases:
The increased ease and frequency of global travel allow infected persons to efficiently spread an infection to a new area.
Rapid urbanization in the major cities of the tropics has created slums characterized by extreme density of human populations.
The profusion of nonbiodegradable containers (i.e., mostly plastic) has created breeding sites for many mosquitoes, especially Aedes aegypti, in proximity to large human populations.
The public health infrastructure has deteriorated, and vector-control programs have been abandoned, due in part to the pressures to “privatize” the economies of many developing countries.
In addition to these proximate causes for the emergence of arthropod-borne viruses, several other factors may have played a role in their emergence in the past or could become more important in the future:
Building of large dams and water projects that provide breeding sites for mosquito vectors
Deforestation and changes in land use associated with expansion of human habitation
Introduction of new virus-amplifying hosts or efficient vectors into new areas (e.g., introduction of Aedes albopictus into the Americas)
Also, climatic changes (global warming) could amplify endemic transmission by expanding mosquito breeding sites to new temperate areas and potentially decreasing the extrinsic incubation period of the virus in the mosquito.4
ARTHROPOD-BORNE VIRUS INFECTIONS
Arboviruses are important causes of encephalitis and hemorrhagic fever in many parts of the world. Only specific species of mosquitoes or ticks that are present in specific ecologic systems can transmit these viruses. Because they depend on transmission by the arthropod host, these diseases are seasonal in temperate climates, yet occur on a year-round basis in tropical climates. The virus must replicate in the vector and travel from the mosquitoes’ stomach to their salivary glands before transmission can occur. The interval between infection of the vector when blood is ingested from a viremic host until the virus appears in the salivary gland of the arthropod is referred to as the extrinsic incubation period. This period is shorter at higher temperature, but averages 6-10 days for most arboviral infections.
Arboviruses are classified into four families: Togaviridae, Flaviviridae, Bunyaviridae, and Reoviridae. The vectors, animal reservoir, and geographic distribution of some important representatives of these four viral families are shown in Table 25-1. The viruses generally are not cytopathic to the infected mosquitoes. However, some arboviruses are neurotropic in the mosquito, as well as in humans. This factor is especially true for Japanese encephalitis (JE) virus and may increase transmission by enhancing biting of CO2-emitting targets, such as humans and other animals, by JE-infected mosquitoes.
The transmission of these viruses may occur throughout the year in tropical areas but increases during the rainy seasons. In contrast, in temperate climates, infections occur only during the warmer months. The mechanism of survival of arboviruses over the winter in cold temperatures is of considerable epidemiologic interest. Although it is possible that a female mosquito could hibernate after a blood meal and reemerge the following season, this behavior is believed to be uncommon. Generally, only nulliparous mosquitoes hibernate. Nevertheless, hibernating mosquitoes infected with West Nile virus have been found in storm sewers during the winter in the New York area.5 The virus could also be reintroduced by migrating birds. In southern areas in the United States, virus-infected mosquitoes have been found year-round.6 Another mechanism for wintering over is the vertical transmission of the viral genome to the eggs of an infected female mosquito. This pattern has been shown to occur among most of the viruses in the Bunyavirus and Phlebovirus groups. Also, experimental evidence indicates that vertical transmission of several flaviviruses is possible, as is sexual transmission of these viruses from male to female mosquitoes.7, 8 Ticks do survive for several years and can spread infection during subsequent seasons after they are infected.
FLAVIVIRUSES
The Flaviviridae family contains approximately 68 single-stranded RNA viruses, most—but not all— of which are transmitted by arthropods. Several flaviviruses have been especially prominent among the important emerging human infections in the past decade or two. Notably, West Nile virus has spread dramatically in the Western Hemisphere, dengue in the Caribbean and Central and South America, and Japanese encephalitis virus in Asia.9
DENGUE AND OTHER MOSQUITO-BORNE INFECTIONS
Although dengue has been known as a human disease for more than 200 years, in the last 20 years, dengue fever, dengue hemorrhagic fever (DHF), and dengue shock syndrome (DSS) have emerged as the most important arthropod-borne viral disease of humans worldwide.10 It is estimated that as many as 100 million cases of dengue fever occur annually on a global basis. Approximately 250,000 cases of DHF are officially reported each year, but the actual number is probably several times higher. The syndrome of DHF/DSS was first reported as an epidemic disease in the Philippines in 1954, and it gradually spread to other areas in Southeast Asia.11 This more severe form of dengue is associated with a mortality rate of 5% to 15% and is believed to be caused by immune complexes formed when a person with a history of one dengue serotype becomes infected with a different serotype.12 The robust immune response is not specific enough to prevent disease, but is strong enough to cause vascular damage by creating immune complexes on FC receptors of vascular endothelial cells. The vascular leakage from the damaged vessels can then cause hemorrhage and fluid accumulation, leading to DHF or DSS.
Immune enhancement was suggested as the cause of DHF in the 1997 dengue 2 outbreak in Cuba.13 That country had been free of dengue since the outbreak of dengue 1 infection in 1981. Overall, it was estimated that 5208 cases of dengue fever and 205 cases of DHF occurred. In sharp contrast to outbreaks in Asia, nearly all dengue fever and DHF cases in this outbreak arose in adults; all except three of the DHF cases were in adults who had been infected with dengue 1 in 1981 and experienced a secondary dengue infection in 1997.14, 15 In another outbreak of dengue in Rayong, Thailand, in 1980, researchers found that all cases of DHF had a secondary immune response, indicating they had had a prior dengue infection.16
In contrast, an outbreak of dengue 2 infection occurred in Peru in 1993 in a population that had previously been infected with other dengue genotypes; however, no cases of DHF occurred.17 This outbreak supports the second major hypothesis concerning the cause of DHF/DSS—namely, that the virulence of the dengue strain is a critical factor in the pathogenesis of DHF and more severe dengue infections. This hypothesis has been put forward by Rosen and colleagues, who have reported DHF in patients during a primary dengue infection.18, 19 and 20 The pathogenesis of DHF/DSS is not completely understood, but various data favor both immune enhancement from a secondary infection and high virulence of some dengue viruses and the combination of these factors to be related to severe life-threatening dengue infections, meaning DHF and DSS.21
e Several other Bunyaviridae (Arbia and Toseana viruses of genus Phlebtovirus; Erve virus of genus Nairovirus), Reoviridae (Lipovnik and Tribec viruses of genus Orbivirus; Eyach of genus Coltivirus), and single tick-borne members of Bunyaviridae (Bhyanjavirus) and Orthomyxoviridae (Thogotovirus) have been tentatively associated with CNS disease in Europe, Asia, and Africa (Dobler, 1996).
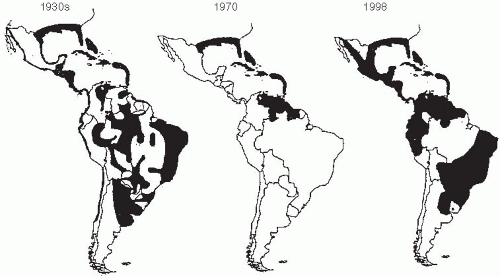
Several factors have promoted the emergence of dengue and its spread to new geographic areas. Increases in the use and disposal of nonbiodegradable containers and their storage in peridomestic locations have provided a wealth of breeding places for the mosquito vector Aedes aegypti in close proximity to humans. A program to eradicate A. aegypti from the Americas was initiated by the Pan American Health Organization (PAHO) in 1947 to control yellow fever, which is also spread by the same mosquito. Efforts were successful in a number of countries. By 1972, A. aegypti had been eliminated from 19 countries in the Americas, representing 73% of the area originally infected (Figure 25-1).
Unfortunately, the campaign gradually lost steam, and funding was withdrawn.22 This occurred in part because of the identification of a jungle cycle of yellow fever and the recognition that, even with the elimination of A. aegypti from urban areas, a focus of yellow fever would persist. Hence, the disease was no longer felt to be eradicable. Within about 10 years, A. aegypti had reestablished itself in virtually all of South and Central America (Figure 25-2).
Figure 25-1 Aedes aegypti distribution in the Americas during the 1930s and in 1970 and 1998. Gubler D. Dengue and dengue hemorrhagic fever. Clin Microbiol Rev. 1998 July; 11(3);480-96. Reproduced with permission from American Society for Microbiology.
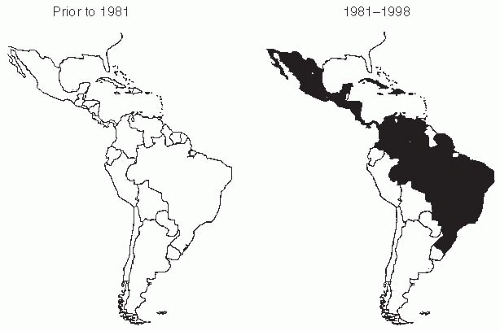
In 1981, a severe epidemic of dengue type 2 infection, including the first cases of DHF/DSS in the Western Hemisphere, occurred in Cuba.23 In 1986, an explosive outbreak of dengue type 1, involving more than 1 million cases, occurred in Rio de Janeiro. Subsequently, dengue epidemics were observed in Paraguay, Bolivia, Peru, Ecuador, Colombia, and Venezuela. More recently, dengue outbreaks have been reported for the first time in Argentina.24 All four dengue serotypes are currently endemic in Latin America. Their continued importance was evident when a large epidemic of dengue type 2 virus, including cases of DHF, occurred in Cuba in 1997.25, 26 and 27
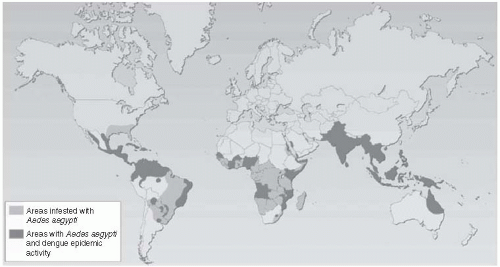
Reports of dengue in Africa were unusual in the 1960s and 1970s or prior to that time. In the mid-1980s, however, dengue appeared first along the coast of Kenya, then in the Ivory Coast and Burkina Faso. Dengue has spread to other African countries in recent years. Also, in Asia, dengue has spread westward into India, Pakistan, and the Middle East in the last decade (Figure 25-3).
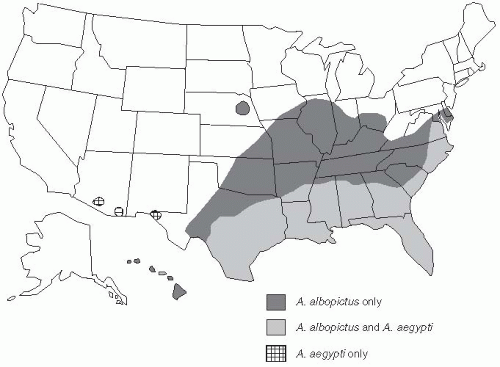
Another factor that has raised considerable concern about the potential for further spread of mosquito-borne viruses in the Americas was the introduction of Aedes albopictus into Houston in used truck tires imported from Southeast Asia for recapping in 1985.28 These spare tires collected rainwater and served as excellent breeding sites for these mosquitoes, which could transmit dengue and yellow fever, as well as other arthropod viruses, such as eastern equine encephalitis. A. albopictus mosquitoes have also been introduced into Brazil in a similar manner; these effective vector mosquitoes are now widely distributed in 19 states of the United States (Figure 25-4). Each year, more than 100 cases of dengue are reported in persons in the United States who have acquired their infections in the endemic areas of Asia, Latin America, or Africa.29 It is quite possible that these viremic individuals might eventually provide a sufficient reservoir to reestablish dengue as an endemic disease in the southern United States, with either A. aegypti or A. albopictus as the vector.
Figure 25-2 DHF in the Americas before 1981 and from 1981 to 1998. Gubler D. Dengue and dengue hemorrhagic fever. Clin Microbiol Rev. 1998 July; 11(3);480-96. Reproduced with permission from American Society for Microbiology.
The prevention of dengue in areas where the disease is endemic is directed at avoiding contact with an infected A. aegypti or A. albopictus vector. Dengue epidemics tend to be geographically localized where both A. aegypti and infected humans reside. An infected mosquito remains infected for its entire life and may bite repeatedly, so it can be a highly effective vector. Because these mosquitoes are found both around homes (peridomestic) and in homes, and because they may bite at any time, especially during the daytime, prevention is difficult. The most effective means of prevention involves destroying sites where larvae develop, such as water-filled containers in and around the house, and using strategies to prevent larval development when water-filled containers are present. Outdoor spraying to kill adult mosquitoes is much less effective than attacking the larval breeding sites, as adult A. aegypti may be indoors when the spraying is done.30
Figure 25-4 A. aegypti and A. albopictus distribution in the United States in 1998. Gubler D. Dengue and dengue hemorrhagic fever. Clin Microbiol Rev. 1998 July; 11(3);480-96. Reproduced with permission from American Society for Microbiology.
While it is possible to control dengue by eliminating larval breeding sites,31 this approach requires a very labor-intensive effort. Household screens, airconditioning, and other methods to seal the living area from mosquitoes are also effective in preventing dengue.
A recent report described a coordinated program to control dengue in several communities in Vietnam.32 This program involved support from the central government (vertical) plus local community action (horizontal approach). It focused on destroying larval breeding sites where possible and the use of predacious copepods of the genus Mesocyclops as a biological agent to control A. aegypti larval development in water-filled containers. The program has eliminated dengue from the target area for the past two years.
Several researchers have been working for more than a decade on developing an effective dengue vaccine.33, 34 and 35 One problem in development of a vaccine to prevent dengue is that the vaccine needs to be equally effective against all four serotypes and protection needs to last until dengue is no longer endemic in an area. If either of these criteria is not met, use of a vaccine could potentially increase the risk of DHF or DSS when the protective immunity wanes. This theoretical risk is based on the hypothesis that DHF/DSS could occur when the level of protective antibodies wanes over time. Tetravalent dengue vaccines have been developed and are currently undergoing clinical trials in humans.
WEST NILE VIRUS IN NORTH AMERICA
West Nile virus (WNV) is a flavivirus that was first isolated in 1937 from the blood of a febrile patient in the West Nile district of northern Uganda.36 Subsequently, in the 1950s, the virus was recognized as a cause of severe meningoencephalitis in the elderly. Equine disease due to WNV was reported in the 1960s in Egypt and France.37 Outbreaks of WNV infection were reported in Israel, France, Russia, South Africa, and Romania in the 1970s.38, 39 Studies of these outbreaks found that birds were the reservoir and amplifying host, that infections were transmitted by mosquitoes, and that horses, birds, and humans were susceptible.40
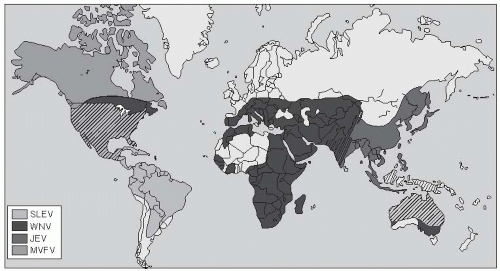
During 1994-2000, many outbreaks of WNV occurred in North America, Europe, and the Middle East as the geographic area for endemic WNV expanded.40 The virus is now recognized to have an extensive distribution in Africa, the Middle East, Eastern Europe, the former Soviet Union, and South Asia, as well as in Australia, where it is known as Kunjin virus (Figure 25-5).40
West Nile virus is antigenically related to Japanese encephalitis virus, St. Louis encephalitis virus, and Murray Valley fever virus. However, unlike the other members of the Japanese encephalitis serogroup, WNV can be divided into two lineages. Lineage 1 viruses are more pathogenic for humans, whereas lineage 2 viruses cause only mild human disease or no symptoms. Lineage 1 viruses have circulated recently in Romania (1996), Tunisia (1997), and Russia (1999) and are now epidemic in the United States and Canada.
Probably the best recent example of the introduction and rapid emergence of a human mosquito-borne viral infection from one geographic region to another is the emergence and spread of West Nile virus in North America after its introduction in the summer of 1999 into the borough of Queens, New York.41 Subsequent genetic analysis work has shown that the genetic structures of the North American WNV isolates and those from Israel are virtually identical.42
The original outbreak was recognized and reported by an infectious diseases physician, who cared for six patients with encephalitis in August 1999.41 He suspected an outbreak of viral encephalitis based on the patients’ similar clinical presentation. Initial serological investigation of the cases detected immunoglobulin M (IgM) antibodies to viruses in the St. Louis encephalitis group of flaviviruses. In fact, initially the outbreak was thought to be due to St. Louis encephalitis (SLE) virus, as SLE had been endemic in North America, although it had never been reported from New York. Subsequently, 62 cases of meningoencephalitis, with 8 deaths, were reported in the New York area in 1999.
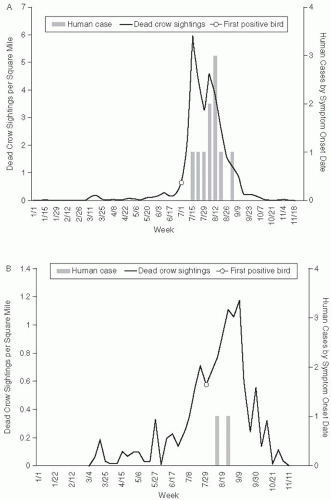
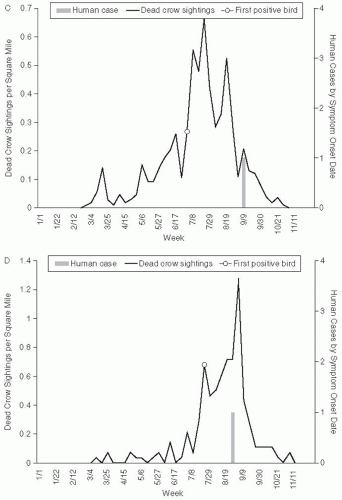
Another unusual feature of this epidemic was the deaths of several bird species at the Bronx zoo in September 1999. Deaths occurred in a cormorant, two Chilean flamingoes, and an Asian pheasant. In addition to the deaths of birds in the zoo, the 1999 outbreak and subsequent outbreaks of WNV were accompanied by extensive mortality among wild birds, especially crows and other corvid species. The extensive bird mortality associated with WNV infection has differentiated this virus from other arbovirus epidemics. In the 2002 WNV epidemic, a total of 124,854 dead birds were reported to state and local health departments, 31,514 were tested for WNV, and 15,745 (50%) were positive. West Nile virus has been isolated from 138 species of birds in the United States.43 In fact, bird mortality has proved a sensitive method of epidemiologic surveillance of the geographic extension of WNV activity in each of the annual epidemics since 1999.44, 45 Bird mortality has generally preceded cases of WNV infection in humans and has been used for sentinel surveillance of WNV activity (Figure 25-6). In addition, in common with several other North American arboviruses, WNV infection is commonly fatal for horses. The impact of WNV emergence in other species is an evolving story. The near-elimination of corvid species from areas of the United States will alter the distribution of many other bird species. Ornithologists are actively working to determine susceptibility to WNV and minimize morbidity and mortality among endangered birds, such as the whooping crane.
Figure 25-6 Dead crow density (number of dead crow sightings per square mile) compared with number of human cases, by week. A. Staten island, axis scale for weekly dead crow density 0 to 7; B. Brooklyn, axis scale for weekly dead crow density 0 to 1.4; C. Queens, axis for weekly dead crow density 0 to 0.7; D. Manhattan, axis scale for weekly dead crow density 0 to 1.4. Reproduced from Eidson et al. Dead crow densities and human cases of West Nile virus, New York State, 2000. Emerg Infect Dis. 2001;7:662-4.
Figure 25-6 (Continued)
After the localized epidemic in New York in the summer and fall of 1999, seasonal epidemics spread to other areas of the United States in subsequent years. In 2001, 64 cases of WNV meningoencephalitis were reported, but the cases were in 38 counties in 10 states in the eastern United States. In 2002, a major epidemic of WNV occurred, in which 4156 cases and 284 deaths were reported. The human cases in 2002 were concentrated in several midwestern (i.e., Michigan, Illinois, and Ohio) and southern states (i.e., Louisiana, Texas, and Mississippi).46 In the summer and fall of 2003, another large epidemic occurred that was especially severe in several western states, including Nebraska, Colorado, North and South Dakota, and New Mexico.47 Overall, 9862 symptomatic human WNV infections and 264 deaths were reported in 2003 (Table 25-2). In 2004, the epidemic was centered in Arizona and California, and 2470 cases and 88 deaths were reported.48 It is somewhat curious that although WNV has spread to Central America and the Caribbean, encephalitis is less frequent there than in North America.
Table 25-2 Species of mosquito positive for WNV by year
 Early History of Infectious Disease: Epidemiology and Control of Infectious Diseases
Early History of Infectious Disease: Epidemiology and Control of Infectious Diseases



