Objectives
- 1.
Map out an overview of meiosis.
- 2.
Describe the general anatomic components of the male and female reproductive systems.
- 3.
Describe the development of the male and female reproductive systems in utero.
- 4.
Describe the regulation of puberty.
- 5.
Describe the changes in adolescent boys and girls that occur during puberty (Tanner stages).
- 6.
Explain the causes and physiologic changes that occur during menopause.
- 7.
Explain the decline of androgens in men (andropause).
Many of the functional deficiencies and clinical syndromes associated with reproduction are linked to the following elements in some way (as a cause or as a consequence):
- •
Embryonic development of the male and female systems
- •
Onset of reproductive maturity and activity at puberty
- •
Age-related changes that lead to a decline in reproductive function with aging ( menopause and andropause )
Thus before discussing the male and female reproductive systems in detail, it is useful to familiarize oneself with the general components of each system and their changes in development and function during life.
General Components of a Reproductive System
The two anatomic components of the reproductive system are the gonads and the reproductive tracts . The gonads ( testes in men, ovaries in women) have both endocrine and exocrine functions, because they produce hormones and gametes (sperm and eggs). The major hormone product of the testes is testosterone . The major hormone products of the ovaries are estrogen before ovulation and progesterone plus estrogen after ovulation. Like the thyroid and adrenal glands, adult gonadal endocrine function is regulated within a hypothalamus-pituitary-gonadal axis . The female reproductive axis is remarkable in that it involves both negative and positive feedback. The testes produce sperm essentially in a continuous manner, whereas the ovaries produce eggs in a discontinuous manner at a rate of about one egg per month.
The reproductive tracts (also referred to as internal and external genitalia ) serve to transport gametes and, in women, allow for fertilization, implantation, gestation, and labor. Because natural fertilization in humans involves internal insemination of the female tract by the male, the male tract includes an intromittent organ, the penis , and the female tract includes the copulatory organ (and birth canal), the vagina . The external portions of the tracts, called the external genitalia , receive innervation that is associated with sexual excitement and orgasm, thereby reinforcing human sex drive. The mammary glands are an adjunct to the female reproductive system, providing nourishment and immune protection to the newborn and infant. Normal gametogenesis in the gonads, the development and physiology of the male and female reproductive tracts, and postpubertal development of the breasts are absolutely dependent on the endocrine function of the gonads.
Overview of Meiosis
Sexual reproduction in humans requires the production of specialized haploid cells (called gametes) through the process of meiosis ( Fig. 8.1 ) . The male gametes are called sperm and are produced in the testes. The female gametes are called eggs (or ova) and are produced in the ovaries. Sexual reproduction has the advantage of generating new genotypes within the progeny, thereby increasing the genetic diversity of a species. Genetic diversity in humans is advantageous primarily because it dilutes out the dosage and effects of deleterious mutations, although it also has adaptive value. The diversification of genetic material occurs during sexual reproduction at three levels:
- 1.
In humans, the 23 chromosomes from the father are complemented by 23 homologous chromosomes from the mother after fertilization. This generates 46,XX or 46,XY individuals with DNA from two unrelated individuals.
- 2.
Meiosis promotes the random recombination of chromosomes. During production of gametes, the 23 pairs of chromosomal homologs undergo independent assortment during meiosis so that the possible combinations of chromosomes in a haploid gamete equals 2 23 , or 8,400,000 genetically distinct gametes.
- 3.
Before entry into meiosis, chromosomes are replicated, generating sister chromatids that closely adhere to each other along their length. During the first meiotic prophase, homologous chromosomes (now as sister chromatids) pair , forming a bivalent . Before or during pairing, double-stranded breaks are induced in a chromatid, followed by the complex process of crossing-over between homologous chromosomes. This process occurs at two or three sites in each chromosome and introduces genetic recombination , thereby further scrambling the DNA that will reside in a gamete.
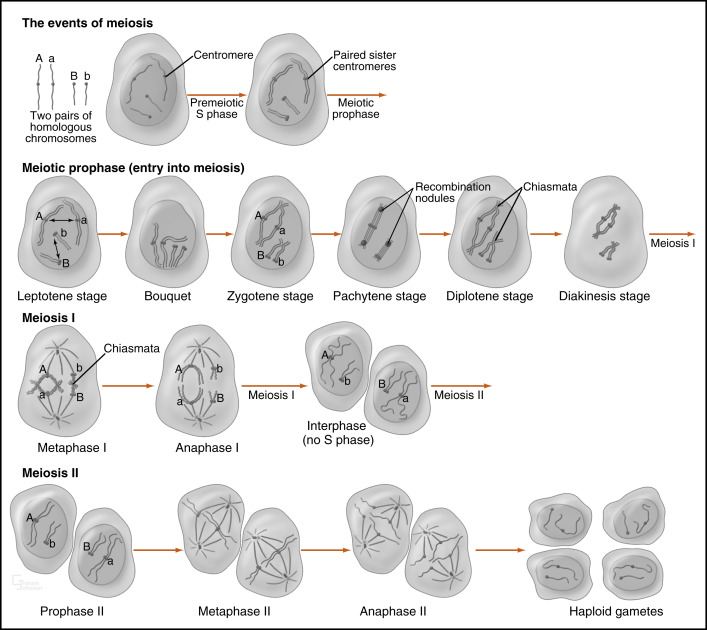
As shown in Fig. 8.1 , meiosis consists of two phases. During the premeiotic prophase, chromosomes are replicated and consist of sister chromatids that are tightly adhered to each other at the centromeres and along their entire length. Prophase I involves the complex process of crossing-over , followed by alignment and disjunction (separation) of the homologous chromosomes. Because of crossing-over, points of chromosomal adhesion, called chiasmata , occur. Chiasmata resist separation of homologous chromosomes and thereby help to guide the alignment of chromosomes along the metaphase plate. Chiasmata are disassembled during anaphase I, allowing the disjunction (separation) of homologous chromosomes.
A kinetochore is a large protein complex associated with the centromere of a chromosome and connects to microtubule spindles that pull the chromatids to opposite poles. Importantly, in meiosis I, the kinetochores of the sister chromatids rotate and fuse to each other. This results in the movement of sister chromatids to the same pole at anaphase I. Consequently, homologous chromosomes are separated during meiosis I, but chromatids are not (see Fig. 8.1 ). After meiosis I, cells enter a brief interphase, but do not replicate DNA (i.e., no S phase). During this time, sister chromatids become separated, except at their centromeres. Meiosis II proceeds much more rapidly and involves the separation of chromatids. This involves the loss of cohesion between sister centromeres at anaphase II so that kinetochores can become attached to spindles on opposite poles. The final product is haploid gametes.
As discussed later, there are striking sex-specific differences in the details of meiotic induction and progression, as well as in gamete maturation, interaction with somatic nurse cells, and release.
Although meiosis and sexual reproduction confer concrete advantages to humans, correct disjunction or separation of chromosomes must occur at both phases of meiosis to maintain the integrity of the genotype (i.e., maintenance of euploidy ). Failure of correct separation in either meiosis I or meiosis II is called nondisjunction and results in the addition of genetic material at one pole and the deletion of genetic material at the opposite pole. Nondisjunction causes aneuploidy (one or more chromosomes are not diploid) or polyploidy (all chromosomes are not diploid). It is likely that the resulting gametes, if they participate in fertilization and embryogenesis, will result in spontaneous abortion. The mistake of nondisjunction occurs with alarming frequency, in that 50% of conceptions are estimated to result in spontaneous abortion, of which more than 60% are caused by aneuploidy. Overall, spontaneous abortion acts as a safeguard against mistakes in chromosome number. In rare cases, aneuploid individuals survive. For example, gain of an extra chromosome 21 ( trisomy 21 ) is compatible with life and results in Down syndrome . These individuals suffer abnormal development of the CNS and a decreased life expectancy.
Basic Anatomy of the Reproductive Systems
Overview of the Male Reproductive System
The testes are the male gonads. Each testis (right and left) resides outside of the abdominopelvic cavity within the scrotum ( Fig. 8.2 ). The testis produces sperm within tubules (seminiferous tubules). The lumina of the tubules empty into the anastomosing network of tubules called the rete testis, which in turn empty into about 20 efferent ductules . Ultimately, efferent ductules converge into one lumen in the epididymis . The right and left epididymides, vas deferens, and ejaculatory ducts transport sperm from each testis to the midline male urethra .
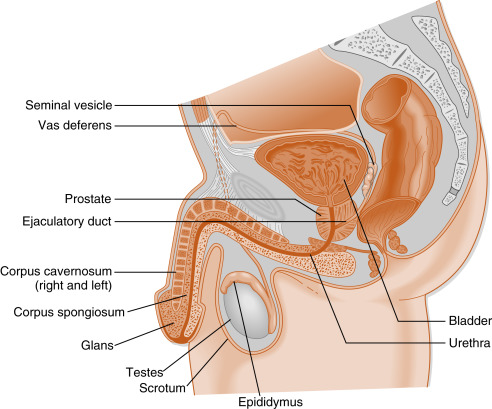
The male tract also contains two large accessory sex glands, the seminal vesicles and the prostate . These glands produce most of the volume of the semen , which mixes with spermatozoa as they enter the urethra. Semen components largely provide a buffered, bacteriostatic, and nutrient-rich microenvironment for sperm after ejaculation into the vagina.
The male urethra becomes the distal portion of the male tract and consists of prostatic (intrapelvic), membranous (a short segment within the deep perineal space), and penile (extrapelvic in superficial perineal space) segments. The urethra receives lubricating and cleansing secretions from the paired bulbourethral glands and the paraurethral glands (glands of Littre). The penile segment courses through the length of the penis. The penis serves as an intromittent organ designed for internal insemination within the vagina. Sensory (pudendal) innervation of the penis leads to orgasm during coitus, thereby reinforcing libido.
The Female Reproductive System
The ovaries are the female gonads. Each ovary (right and left) resides within the pelvic cavity ( Fig. 8.3 ). There are no tubules in the ovary. Instead, the gametes (primary oocytes) become invested with epithelial and stromal cells that make up an ovarian follicle; one follicle contains a single oocyte . With growth, the follicle ultimately creates a fluid-filled lumen (called an antrum ). At the time of gamete release, the gamete (now an egg arrested in meiotic metaphase II) and a thin covering of epithelial cells become free-floating within this lumen. Ovulation involves a complex set of events that essentially erode the follicular and ovarian walls at the point where the follicle is pushing against the ovarian surface, followed by the release of the egg.
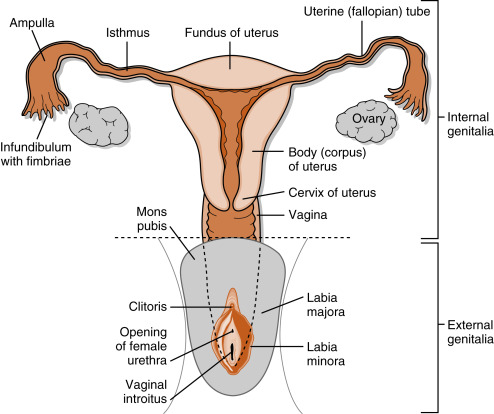
Unlike the male tract, the egg is released into the pelvic cavity and has to be captured by the proximal segment of the female tract called the oviduct (see Fig. 8.3 ). The oviduct has an opening at its proximal, free end (the infundibulum), through which the captured egg is transported. There is usually only one egg ovulated, either from the right or left side, depending on which side had the largest follicle at the beginning of the menstrual cycle. The oviduct transports the egg toward the midline uterus and allows for the movement of sperm from the uterus laterally toward the egg. Fertilization and early development (5 to 6 days) normally occur in the oviduct. The early embryo (blastocyst) eventually moves into the uterine lumen and implants into the uterine mucosa. The growing fetus is supported in part by the elastic and fibrous inferior end of the uterus called the cervix . At term, the newborn is expelled from the uterus through the cervix and vagina.
The vagina acts as both the copulatory organ and the birth canal. The female external genitalia surround the superficial opening of the vagina (called introitus ). The labia majora are homologous to an unclosed scrotum. Vestibular bulbs (deep to the labia minora ) and the clitoris represent structures homologous to the erectile tissue of the penis. However, unlike the penis, erection of these structures is not required for fertility. Sensory (pudendal) innervation of these structures and the vaginal wall may lead to orgasm during coitus, thereby providing reinforcement to libido.
Sexual Development in Utero
The genetic sex of a fetus depends on the nature of the sex chromosomes contributed by the egg and the sperm. Normally, there are 46 chromosomes, consisting of 22 pairs of autosomes and one pair of sex chromosomes. The sex chromosomes are called X and Y chromosomes; 46,XX is the normal karyotype for the female, and 46,XY is the normal karyotype for the male. Genetic sex determines gonadal sex . The gonads then either produce hormones (if male) or no hormones (if female). The hormonal environment determines the sex of the reproductive tract and external genitalia.
Male Development
During the first 6 weeks of development, mesodermal cells within the genital ridge develop into a bipotential gonadal primordium ( Table 8.1 ). During this period, primordial germ cells migrate from the yolk sac endoderm into the gonadal tissue. By 6 weeks of gestation, the gonads contain germ cells, supporting cells, and stromal cells that will become androgen-producing steroidogenic cells in both sexes.
| Gestational Age (wk) | Developmental Feature |
|---|---|
| 6-8 | Differentiation of testes |
| 8 | Retention of Wolffian ducts |
| Regression of Müllerian ducts | |
| 9-12 | Development of male-type external genitalia |
The short arm of the Y chromosome contains the SRY gene (sex-determining region on the Y chromosome) that encodes the transcription factor, SRY . SRY, along with other transcription factors, plays a major role in the induction of differentiation of the bipotential gonad into a testis between weeks 6 and 7 of gestation ( Fig. 8.4 ). With testicular development, the supportive epithelial cells differentiate into Sertoli cells and form the seminiferous tubules. Sertoli cells perform two critical functions in early embryogenesis: (1) they surround male germ cells (called spermatogonia ) and produce an enzyme (CYP26B1; also produced by Leydig cells) that degrades locally produced retinoic acid , thereby preventing meiotic progression of spermatogonia; and (2) they express antimüllerian hormone (AMH , also referred to as müllerian-inhibitory substance ), which causes the degeneration of the müllerian ducts (see later in the text).
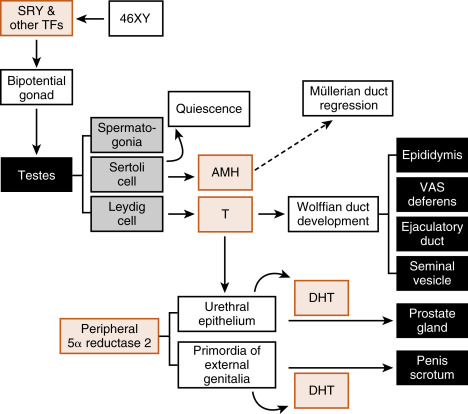
Hormones mediate phenotypic gender expression. The fetus originally develops with bipotential internal and external genitalia ( Figs. 8.5 and 8.6 ). Internally, there are two Wolffian (also called mesonephric ) ducts , which have the potential for differentiating into the nonurethral segment of the male tract (epididymis, vas deferens, ejaculatory duct, and seminal vesicles), and two Müllerian (also called paramesonephric ) ducts , which have the potential for differentiating into most of the female reproductive tract (the oviducts, uterus, cervix, and proximal third of the vagina). Whether male or female reproductive tracts develop depends on the presence or absence of two hormones produced by the fetal testis- testosterone and AMH ( Box 8.1 ).

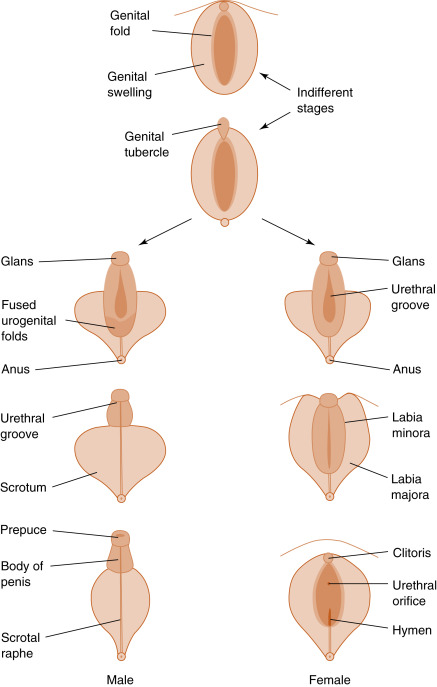
- •
The Wolffian ducts, when stimulated with testosterone, become the epididymis, vas deferens, seminal vesicles, and ejaculatory ducts.
- •
The Müllerian ducts, in the absence of Sertoli cell Müllerian-inhibiting substance, become the fallopian tubes, uterus, cervix, and upper one-third of the vagina.
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


