Table 58.1 Immunochemical and enzymatic markers of coagulation and fibrinolytic system activation | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
thrombomodulin on vascular endothelial cells and once bound can no longer participate in procoagulant mechanisms, either to activate platelets or to cleave fibrinogen. Endothelial protein C receptor, which lies adjacent to thrombomodulin within the caveolae region of the endothelial membrane, augments this mechanism by capture of protein C from the circulation, particularly in large vessels, and presents protein C to the thrombin-thrombomodulin complex. Thrombin-thrombomodulin activates protein C, which then inactivates factor Va and factor VIIIa by cleavage, thereby also regulating thrombin generation. Assays have been developed to monitor this pathway (FIGURE 58.2). Immunoassays to measure protein C activation include measurements of the protein C activation peptide,24 an immunoactivity assay for activated protein C,56,57,58 and activated protein C-inhibitor complexes.26,27,59
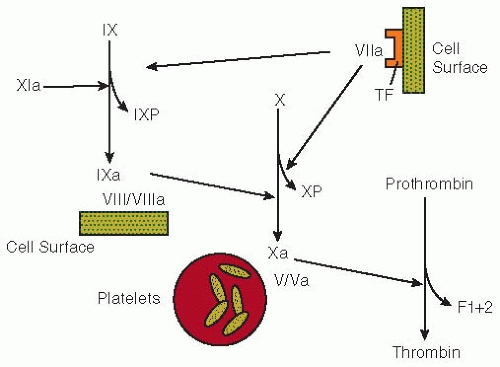 FIGURE 58.1 Pathways of coagulation activation. The activation of factor IX by factor XIa or the factor VII-tissue factor (TF) mechanism liberates the factor IX activation peptide (IXP). The conversion of factor X to factor Xa by the factor IXa-factor VIII/VIIIa-cell surface complex releases the factor X activation peptide (XP). The generation of thrombin from prothrombin is mediated by factor Xa in the presence of factor Va and activated platelets. During this process, the F1+2 fragment is released. |
the smallest. This fibrin derivative is comprised of two fragment D moieties covalently linked by their γ chains.80,81 D-dimer levels in whole blood can be measured by red cell agglutination, whereas levels in plasma can be quantified using an enzyme immunoassay (EIA) or latex bead agglutination. All assays use monoclonal antibodies that recognize neoepitopes on D-dimer that are not expressed on the D-domains of non-cross-linked fibrinogen or fibrin.44,45 Although agglutination assays can be done more rapidly than the EIA, latex agglutination assays are less sensitive and provide only semiquantitative results.
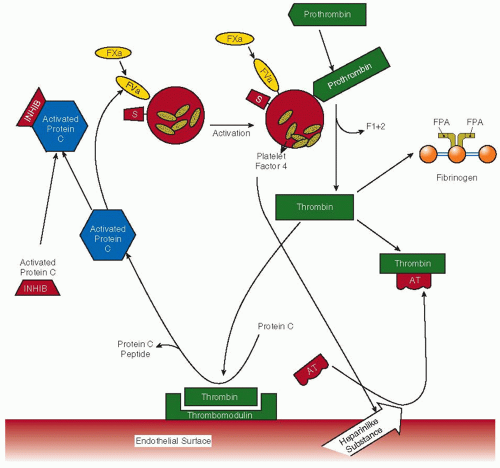 FIGURE 58.2 Regulation of thrombin generation by the natural anticoagulant mechanisms of the endothelium. Factor Xa, factor Va, protein S, prothrombin activation fragment F1+2, fibrinopeptide A, antithrombin, and activated protein C inhibitor(s) are designated as FXa, FVa, S, F1+2, FPA, AT, and activated protein C inhibitor, respectively. Thrombin may be inactivated by forming 1:1 stoichiometric complexes with its major physiologic inhibitor, antithrombin, thereby resulting in the formation of TAT complexes. Activated protein C can be neutralized by inhibitors of activated protein C (e.g., protein C inhibitor, α1-proteinase inhibitor, and α2-macroglobulin), which results in the generation of activated protein C inhibitor complexes. |
thrombus.85 In addition, D-dimer levels can be elevated in the absence of concomitant evidence of angiographic reperfusion after thrombolysis.85 There are two potential explanations for these findings. The first is that a flaw in the design of D-dimer assays can limit its specificity and lead to higher than expected plasma levels.64,86 Although a monoclonal antibody is used to “capture” the D-dimer in plasma, a panspecific antibody is sometimes used to identify the bound antigen. Cross-reactivity of this “tag” antibody with FDPs would lead to an overestimate of D-dimer levels in the setting of thrombolytic therapy. Rather than reflecting clot lysis, a second possibility is that the elevated D-dimer levels after thrombolytic therapy may reflect large amounts of circulating soluble cross-linked fibrin,64 thereby explaining the increased D-dimer levels that occur in the absence of angiographic evidence of reperfusion.64
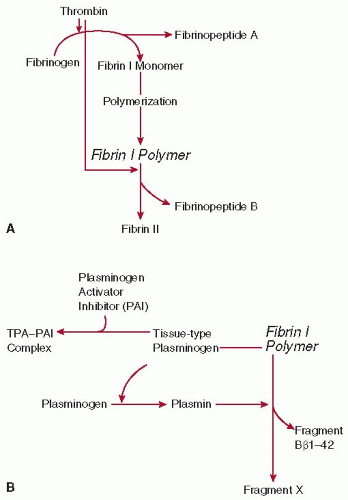 FIGURE 58.3 The actions of thrombin (A) and plasmin (B) on fibrin I polymer. A: The conversion of fibrinogen to fibrin II polymer by thrombin. B: The action of plasmin on fibrin I polymer. The fibrinolytic capacity of human plasma is determined primarily by the activity of tissue-type plasminogen activator (tPA) and its inhibitor (plasminogen activator inhibitor 1 [PAI-1]), which are released from vascular endothelial cells. tPA is able to bind to fibrin I polymer, which allows plasminogen to be activated to plasmin at an increased rate. |
Related posts:
 The Field of Hemostasis and Thrombosis: Selected Translational Achievements
The Field of Hemostasis and Thrombosis: Selected Translational Achievements
 Structure and Function of Vitamin K-Dependent Coagulant and Anticoagulant Proteins
Structure and Function of Vitamin K-Dependent Coagulant and Anticoagulant Proteins
 Integrín αIIbβ3 and Platelet Aggregation
Integrín αIIbβ3 and Platelet Aggregation
 Acquired Nonimmune Thrombocytopenia
Acquired Nonimmune Thrombocytopenia
 Pathogenesis and Treatment of Biomaterial-Associated Thrombosis
Pathogenesis and Treatment of Biomaterial-Associated Thrombosis
 Blood Management in the Cardiovascular Surgical Patient
Blood Management in the Cardiovascular Surgical Patient
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


