Hepatitis viruses and hepatoma
Hongyang Wang, MD  Guangwen Cao, MD, PhD
Guangwen Cao, MD, PhD  Jing Fu, MD, PhD
Jing Fu, MD, PhD  Guishuai Lv, MD
Guishuai Lv, MD
Overview
Hepatitis is an inflammation of the liver, occurring as the result of a viral infection or the exposure of liver to toxic substances such as alcohol or aflatoxin B. Hepatitis viruses are the most common cause of hepatitis in the world, of which HBV and HCV can cause persistent liver infection, frequently resulting in chronic hepatitis, liver cirrhosis, and eventually hepatocellular carcinoma (HCC). HBV belongs to the genus Orthohepadnavirus of the Hepadnaviridae with a circular incomplete double-stranded DNA genome, containing four overlapping open reading frames (ORFs) that encode the surface envelope protein (HBsAg), the core protein (HBcAg and HBeAg), a polymerase, and a multifunctional nonstructural protein termed X (HBx), respectively. HBV genotypes and subgenotypes have distinct geographical distributions. In East Asia, HBV genotypes B and C are endemic. Accordingly, any differences in its global incidence may be explained by differences in the HCV and HBV prevalence. It has been estimated that 57% of cirrhosis is attributable to either HBV (30%) or HCV (27%) and 78% of HCC is attributable to HBV (53%) or HCV (25%). In China, up to 80% of HCC cases are attributable to HBV, and approximately 20% of HCC patients test positive for HCV-RNA. Besides the oncogenic function of HBV-encoding protein, the HBV infection-induced virus or host DNA mutation has been found contribution for HCC development. Unlike to HBV, HCV belongs to the genus Hepacivirus of the Flaviviridae family, with a single-stranded positive-sense RNA genome. There is no host genome integration, nor does HCV contain any known oncogenes. The major mechanisms of HCV-induced hepatocarcinogenesis include the oncogenic effect of HCV viral proteins, steatosis and insulin resistance, chronic inflammation and fibrosis, oxidative stress, and chromosomal instability. This chapter exclusively focuses on hepatitis and hepatoma caused mainly by HBV and HCV infection, discussing epidemiologic considerations, construction and genotype, role of virus in performing HCC, and early diagnosis and prophylaxis. Still, viruses that selectively infect endemic host predispose patients to malignant hepatoma, to some extent, placing a burden on the healthcare worldwide. Therefore, an upturn in morbidity of hepatitis and hepatoma will be of great importance in public health.
Hepatitis and hepatoma
Liver diseases, including hepatitis B virus (HBV) and hepatitis C virus (HCV) infections, alcoholic liver disease (ALD), nonalcoholic fatty liver disease (NAFLD) and associated cirrhosis, and HCC, are major causes of illness and death worldwide.1 Liver disease causes serious public health problems because of its high prevalence worldwide and poor long-term clinical outcome, including premature deaths from liver decompensation, cirrhosis, and HCC. Among liver diseases, hepatitis and HCC harm to people’s health seriously due to the disease susceptibility, cryptobiosis, and refractoriness, respectively.1
Human hepatitis viruses
Human viruses that selectively infect human hepatocytes and cause liver diseases are termed as human hepatitis viruses including hepatitis A virus (HAV, HBV, HCV, hepatitis D virus (HDV), and hepatitis E virus (HEV). Through fecal-to-mouth transmission, HAV or HEV causes temporary hepatic infection, varying from an asymptomatic infection to a fulminant disease. HAV, a single-stranded RNA virus, is a major cause of acute viral hepatitis worldwide. It occasionally causes an acute liver failure. HEV, a single-stranded, non-enveloped RNA virus, is endemic in several Asian and African countries. Pregnant women and patients with pre-existing chronic liver diseases are at a high risk of fulminant liver failure upon HEV infection.2 HBV (alone or co-infected with HDV or super-infected with HDV) and HCV can cause persistent liver infection, frequently resulting in chronic hepatitis, liver cirrhosis, and eventually HCC.
Hepatitis B virus
HBV belongs to the genus Orthohepadnavirus of the Hepadnaviridae with a circular incomplete double-stranded DNA genome of about 3.2 kb in length. HBV genome contains four overlapping ORFs that encode the surface envelope protein (HBsAg), the core protein (HBcAg and HBeAg), a polymerase, and a multifunctional nonstructural (NS) protein termed X (HBx). The PreS region that consists of PreS1 (nt.2848-3204) and PreS2 (nt.3205-154) domains overlaps a region encoding the polymerase gene (P gene). The enhancer II (EnhII; nt.1636-1744) and basic core promoter (BCP; nt.1751-1769) regions overlap with the X gene (nt.1374-1835). HBV genome contains two viral enhancers that regulate transcription of the HBV promoters, including the BCP that controls the transcription of the pre-core and core regions. Following the entry into hepatocytes, HBV enters the nuclei of hepatocytes. HBV genome forms a relaxed circular DNA that is converted into covalently closed circular DNA (cccDNA) in the nuclei. The cccDNA transcribes all viral RNAs including pregenomic (pg) RNA as a replicative RNA intermediate. The pgRNA, viral core, and polymerase proteins are assembled into the nucleocapsid in the cytoplasm and then the pgRNA is converted into an HBV DNA by viral polymerase. Approximately 1011 viral particles are released into the circulation of the individuals with chronic HBV infection per day, and HBV particles are cleared from the plasma with a half-life of approximately 1.2 day. During viral replication, the partially double-stranded HBV DNA is generated from an intermediate RNA via reverse transcription activity of the viral polymerase. HBV reverse transcriptase lacks proofreading capacity, resulting in mutation rates of 1.5 × 10−5 to 5 × 10−5 nucleotide substitutions/site/year in HBV-infected subjects at their HBeAg-positive stage.3, 4 However, owing to the extreme overlapping ORFs, HBV genome evolution is constrained to maintain essential protein functions required for viral replication.5 This plasticity of HBV genome allows the generation of viral mutants that can occur under the pressures of immune selection.
Eight genotypes (genotypes A–H) have been determined according to a sequence divergence >8% in the entire HBV genome or a sequence divergence >4% in the S region. Genotypes have further been classified into subgenotypes if the divergence in whole nucleotide sequence is between 4% and 8%. Subgenotypes 1–5 of genotype A, subgenotypes 1–8 of genotype B, subgenotypes 1–8 of genotype C, and subgenotypes 1–7 of genotype D have been recently nominated. HBV genotypes and subgenotypes have distinct geographical distributions.3, 6, 7 HBV genotypes A1, A3, A4, and A5 are endemic in Africa, especially in West Africa, whereas genotype A2 is endemic in Europe. Genotypes B and C are endemic in Asia. Of HBV genotypes B and C, subgenotypes B2 and C2 are endemic in most parts of Asia. Subgenotype C4 is encountered in Aborigines from Australia and frequently termed as the Australian aboriginal strain. Subgenotypes B3–B8, C1, C3, and C5–C8 are present in Indonesia and the Philippines. Genotype D is endemic in the entire Old World including Northern Africa, Northern and South Asia, the Mediterranean area, and most European countries. Subgenotype D1 is predominant in Moslem ethnicity. Subgenotype D2 is endemic in Russia and the Baltic region. Subgenotypes D4 and D6 are endemic in Oceania and Indonesia, respectively. HBV genotype E is endemic in Western and Central Africa. HBV genotypes F, G, and H are endemic in Middle and Southern America. HBV genotype and subgenotype correlate not only with the clinical outcomes but also with the response to interferon-α treatment.7, 8 In East Asia where HBV genotypes B and C are endemic, genotype B is more apt to cause acute infection in young people and to be more easily cleared than genotype C, whereas genotype C leads to higher persistence following an acute course and is more apt to cause cirrhosis and HCC than genotype B.8–11 Thus, HBV genotyping is not only important in reconstructing the evolutionary history of HBV and humans but also helpful in indicating clinical outcomes of HBV infection and responses to antiviral treatments.
Chronic HBV infection frequently occurs in individuals infected perinatally or during early childhood, when the immune system is immature. HBV infection in adults is mostly asymptomatic or experiences an acute course. Invasive medical procedures, household contact with HBV carriers, body care and beauty treatments, and lack of HBV vaccination are the major risk factors of acute hepatitis B in adults. About 8.5% of adult patients with acute hepatitis B in mainland China will develop a chronic infection.9 HBV genotype C (vs genotype B) and genotype D (vs other HBV genotypes) are more apt to cause persistent infection following an acute course.9, 11 Genetic polymorphisms of human leukocyte antigen (HLA) in the HLA-DP and HLA-DQ regions contribute to immune imbalance (such as Th1/Th2 cells or Th17/Treg cells, neutrophil/lymphocyte, neutrophil/CD8+ T cell, and Th1/Th2 cytokines balances) upon HBV infection, resulting in HBV persistence and possible chronic liver inflammation.12–14 Allelic frequencies of the HLA-DP polymorphisms differ greatly among races. According to the NCBI database (http://www.ncbi.nlm.nih.gov/projects/SNP/), the HLA-DP (or HLA-DQ) alleles that facilitate chronic HBV infection are the major allele in the Asian population,13, 14 whereas they are the variant alleles in the European population. This might be one of the reasons why HBV persistence is more frequent in Asians than in Europeans. These HLA-II genetic polymorphisms may predispose the host to maintaining chronic HBV infection, facilitate the immune selection of the disease-related HBV mutants, and affect the risks of cirrhosis and HCC contributed by the HBV mutations.
Hepatitis B virus and hepatocellular carcinoma
In general, without efficient treatment, all types of chronic hepatitis will finally progress into ESLDs (end-stage liver diseases), such as cirrhosis and HCC. Most ESLD displays a poor clinical outcome. HCC is a major cause of cancer mortality worldwide, and any differences in its global incidence may be explained by differences in the HCV and HBV prevalence.1 In 2013, the World Health Organization reported that primary liver cancer caused 745,517 deaths worldwide and that HCC represented the major histological type of these liver cancers.15 In another comprehensive review of global mortality, deaths from HCV-related HCC were estimated at 195,700 in 2010. In China, HCC is the second leading cause of cancer mortality, and its annual death rate was 24.15 per 100,000 persons in 2009.16 Approximately 383,203 persons die from liver cancer every year in China, which accounts for 51% of the deaths from liver cancer worldwide.16 Up to 80% of HCC cases in China are attributable to HBV, and approximately 20% of HCC patients test positive for HCV-RNA.17
Epidemiologic considerations
Approximately 2 billion people have ever been exposed to HBV worldwide. Of those, 350 million people are chronically infected with HBV. It has been estimated that 57% of cirrhosis is attributable to either HBV (30%) or HCV (27%) and 78% of HCC is attributable to HBV (53%) or HCV (25%).18 A prospective study conducted in Taiwan has proved that the cumulative lifetime (age 30–75 years) incidences of HCC for men and women positive for both HBsAg and antibodies against HCV are 38.35% and 27.40%; for those positive for HBsAg only, 27.38% and 7.99%; and for those positive for neither, 1.55% and 1.03%, respectively.19 Prospective epidemiological studies have proved that male gender, increasing age, cirrhosis, high viral load, HBeAg positivity, HBV genotype C (vs genotype B), low albumin, alanine aminotransferase (ALT) elevation, and viral mutation A1762T/G1764A independently increase HCC risk in chronic HBV-infected patients.11, 19–23 HBV demonstrates “mutation–selection–adaptation,” a viral evolutionary process involved in hepatocarcinogenesis. During this process, especially during HBeAg seroconversion, HBV accumulates HCC-risk mutations, predominantly in the core promoter and preS regions of HBV genome.24–26 A1762T/G1764A can predict HCC prospectively, possibly because this mutation is earlier generated during the evolutionary process.27 These demographic, clinical, and viral factors harvested before HCC occurrence should be prognostic and even predictive for HCC in HBV-infected patients.
Role of HBV mutations in HCC development
Insufficient immune responses elicited by HBV antigens select the HCC-related HBV mutations during long-term evolutionary process. Only the HBV strains/variants best adapted to the immune system will survive and thrive in liver. Genetic predispositions of HLA-II antigens and other inflammatory factors such as NF-κB and STAT3 may contribute to immune imbalance upon HBV infection, leading to persistent infection and chronic inflammation in liver, which facilitate the generation of the HCC-risk HBV mutations.13, 28, 29 Inflammatory factors promote HBV mutations, at least partially, via activating cytidine deaminases.5
Host somatic mutations
Cytidine deaminases and their analogs, whose expressions and activities can be activated by proinflammatory cytokines generated during the inflammation, not only promote HBV mutagenesis but also facilitate somatic mutations.5 Important HCC-related somatic mutations are found in critical genes such as RNA editing genes (ADAR1, ADAR2, KHDRBS2, and RTL1), chromatin remodeling genes (ARID1A, ARID1B, and ARID2), DNA-binding genes (HOXA1), growth factor signaling pathway genes (CDH8, CDK14, CNTN2, ERRFI1, RPS6KA3, P62, and PROKR2), transcriptional regulation genes (AXIN1, CCNG1, CTNNB1, IRF2, NFE2L2, PARP4, PAX5, ST18, TP53, TRRAP, and ZNF717), cell structure modification genes (FLNA and VCAM1), epigenetic modification genes (MLL3), and JAK/STAT pathway genes (JAK1 and JAK2).30 These somatic mutations should affect major signaling pathways, which can serve as therapeutic targets for HCC aggressiveness.
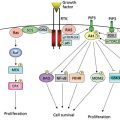
The roles of HBV-encoding proteins
(1) Excess production of the HBV envelope proteins, or truncated forms of the middle (preS2/S) and large (preS1/preS2/S) envelope protein, can activate cellular signal transduction pathways or endoplasmic reticulum (ER) stress pathways, induce cell proliferation by upregulating cyclin A expression, and activate c-Raf-1 and extracellular regulated kinase (ERK) signaling to stimulate cell proliferation. Accumulation of the S proteins in the ER can activate the unfolded protein response and cause oxidative stress. Epidermal growth factor receptor (EGFR) overexpression, which occurs in 40–70% of human HCCs, has been linked with tumorigenesis.31 Aberrant activation of Raf-MEKERK and PI3K-Akt pathways driven by EGFR is commonly observed and implicated in the tumor growth and progression of many human cancer types, including HCC.32 Moreover, activation of EGFR signaling pathways via the high expression of either its cognate ligands or itself is strongly associated with the poor prognosis of HCC. (2) HBx serves as a transactivator of several cellular signaling pathways including Wnt that contributes to HBV-related HCC and also interacts with tumor suppressor adenomatous polyposis coli (APC) to activate Wnt/β-catenin signaling. HBx promotes the invasive ability and metastatic potential of HCC. For example, as recently reported, using HBx transgenic mice and human HBV-related HCC specimens demonstrates that expression of HBx promotes expansion and tumorigenicity of HPCs (hepatic progenitor cells) that contribute to HBx-mediated tumor formation in a diethyldithiocarbamate (DDC)-induced mouse model. These studies shed novel light on the notorious role of HBx in the relationship between chronic hepatitis infection and liver cancer.33
Related posts:
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


