within the submucosa, and has a relatively favorable prognosis compared with EBV-negative GC, although both findings remain to be confirmed.5
Table 4-1 Comparison of H. pylori and the EBV as causative agents of GC | ||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||
differentiated carcinoma with dense infiltration of lymphocytes, resembling NPC.3 LE-like GC is nearly identical to the subgroup that Watanabe et al.7 reported as “GC with lymphoid stroma (GCLS),” but GCLS is relatively a broad category that includes LE-like GC. The relative ratio of the LE-like/GCLS type to the ordinary type reported in the literature varies considerably from 1:10 to 4:1.6 This variability is due to interpretation variability in diagnostic criteria of the LE-like/GCLS type, especially when there is heterogeneity within the tumor. More than 80% of the LElike/GCLS-type tumors were positive for EBV in most reports.3,6,8 The histopathological features of EBV-positive LE-like/GCLS tumors, which are not seen in EBV-negative LE-like/GCLS tumors, are mild cellular pleomorphism, rare mitotic figures, a marked degree of lymphoid stroma, and lymphoid infiltration within the cancer cell nests.8
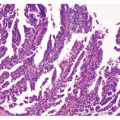
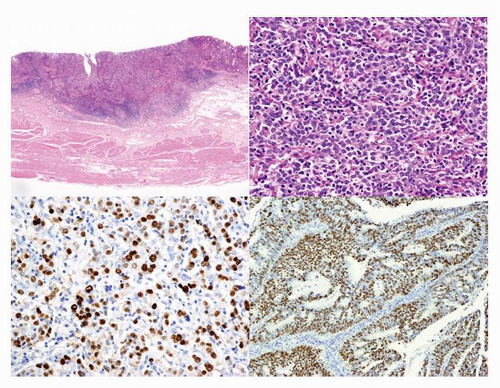 FIGURE 4-1 Histopathology of EBV-associated GC. A (left upper): Low-power view of EBV-associated GC. Lymphoepithelioma (LE)-like carcinoma shows expansive growth with prominent infiltration of lymphocytes. B (right upper): High-power view of LE-type of EBVassociated GC shown in (A). C (left lower): EBER in situ hybridization of LEtype of EBV-associated GC (A). D (right lower): EBER in situ hybridization of ordinary type of EBV-associated GC. |
Table 4-2 Clinicopathological features of EBV-associated GC | ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||
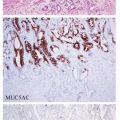
constitute the tight junction, and neoplastic cells of EBV-associated GC showed a high frequency of CLDN18 expression (84%) and a low frequency of CLDN3 expression (5%).10 This expression profile (CLDN18+, 3-) corresponds to that of the normal gastric epithelium in adults and fetuses, but not to that of intestinal metaplasia (CLDN18-, 3+). In accordance with the CLDN expression patterns, almost half of the EBV-associated GC cases in a study by Barua et al.11 had gastric-type mucin expression (MUC5AC, MUC6), and the other half lacked gastric- or intestinal-type mucin or CD10 expression. These results indicate that EBV-associated GC is very homogenous with regard to cellular differentiation and that it preserves the nature of the cells of origin. Thus, the cells that are targets of EBV infection and their subsequent transformation may be precursor cells with intrinsic differentiation potential toward the gastric cell type.
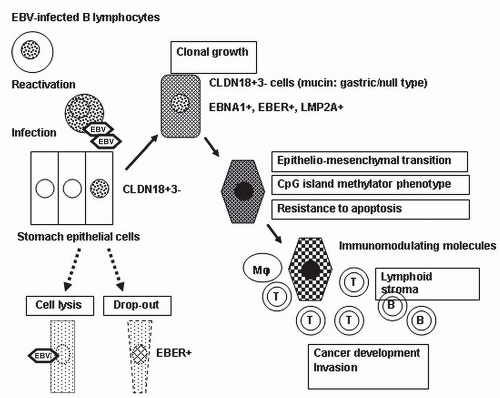 FIGURE 4-2 Schematic representation of the development and progression of EBV-associated GC. Stomach epithelial cells are infected with EBV from the reactivation of EBV-carrying lymphocytes at the mucosa. Some EBV-infected cells begin to grow clonally in the milieu of atrophic gastritis. The neoplastic cells retain the claudin 18-positive/claudin 3-negative (CLDN18+, 3-) gastric phenotype, which may correspond to the phenotype of the original EBV-infected cells. The CpG-island methylation phenotype is the primary abnormality in neoplastic cells of EBV-associated GC. Expression and secretion of immunomodulator molecules induce the characteristic stromal reaction. |
host genome and infection is latent, with no viral replication. Monoclonal or biclonal EBV was observed in carcinoma tissues in the intramucosal stage; however, it was always monoclonal in the submucosal invasion stage (Fig. 4-3) and in more advanced carcinomas.12,13 Since all carcinoma cells show a positive signal in EBER-ISH in all cases of EBV-associated GC, EBV infection must occur at the initial or a very early stage of carcinoma development.
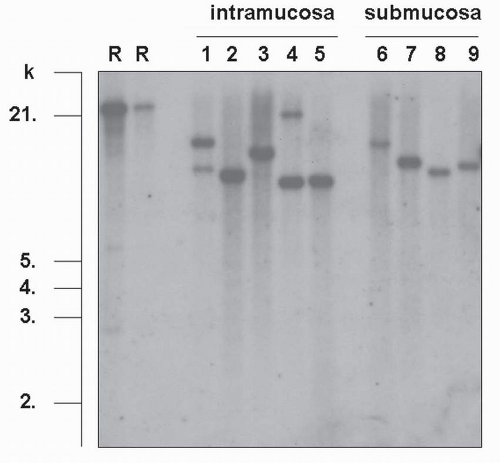 FIGURE 4-3 Clonal analysis of EBV derived from virus-associated GC. Southern blot hybridization analysis of TRs of EBV DNA, extracted from EBV-associated GC. DNA is cut with the BamHI restriction enzyme, electrophoresed, and hybridized with an EBV-TR-specific probe (R, R’: Raji cell used as a control, 1-5: samples of intramucosal carcinomas, 6-9: samples of submucosal carcinomas). Analysis of the blot shows either one or two bands larger than 6 kb in each sample. When the detected bands are short, it indicates that EBV takes the linear form in viral particles. Smeared signals indicate infection of the tumor with various EBV clones. (From Uozaki H, Fukayama M. Int J Clin Exp Pathol. 2008;1:198-216[2], with permission from the Journal.) |
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


