CYTOGENETIC PRINCIPLES
NOMENCLATURE AND IDENTIFICATION OF CHROMOSOMES
Normal humans have 46 chromosomes—44 autosomes and two sex chromosomes. In females, the two sex chromosomes are X chromosomes; in males, one is an X and one a Y chromosome. By convention, chromosomes are numbered according to size and the position of the centromere, the primary constriction around which cell division occurs. As with other chromosomes, the centromere divides the X and the Y chromosomes into a short arm, designated p, and a long arm, designated q. Accepted nomenclature dictates that specific chromosome bands are designated by the chromosome, the arm (p or q), the region, and the specific band; bands are numbered consecutively from the centromere distally. Complements containing structurally abnormal X or Y chromosomes also require the symbol for the aberration present (e.g., i for isochromosome, del for deletion).1,2 Beginning in 1969, the development of banding techniques that produce horizontal “stripes” on the chromosome permitted the routine identification of individual chromosomes1,2a (Fig. 90-1D). Although the mechanism of chromosome banding is not completely understood, the various bands are known to reflect variations in DNA base sequences.
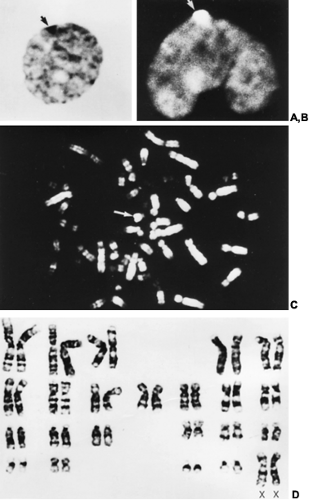 FIGURE 90-1. A, Nucleus of normal female cell from orcein-stained smear of buccal mucosa showing one X-chromatin mass, or Barr body (arrow). B, The nucleus of a peripheral leukocyte from a normal male, stained with quinacrine mustard and showing the fluorescent Y-chromatin (arrow). C, Quinacrine-stained metaphase chromosomes of a normal male from leukocyte culture: arrow indicates fluorescent Y chromosome. D, Giemsa-banded karyotype of normal female from leukocyte culture. Each human chromosome can be identified by its banding pattern. Giemsa banding (G-banding) is the most widely used procedure. The number of bands identified with this technique is ˜500, which permits the detection of subtle chromosome alterations. Additional techniques include quinacrine banding (Q-banding), constitutive heterochromatin banding (C-banding), and reverse banding (R-banding). (Courtesy of Dr. Beverly White; reproduced from Rebar RW. Practical evaluation of hormonal status. In: Yen SSC, Jaffe RB, eds. Reproductive endocrinology: physiology, pathophysiology, and clinical management, 2nd ed. Philadelphia: WB Saunders, 1986:683.) |
CYTOLOGIC PROPERTIES OF THE X CHROMOSOME
One of the two X chromosomes in normal females is the last chromosome to complete DNA synthesis, a fact believed to reflect tighter condensation during interphase of this chromosome than of others. The late-replicating X is said to represent the heterochromatic X and appears as a planoconvex body, now called X-chromatin body and formerly known as sex chromatin or the Barr body, near the nuclear membrane3 (see Fig. 90-1A). These bodies are observed in 30% or more of the cell nuclei from normal females. X-chromatin is the more nonfunctioning X chromosome and is present in the resting nuclei of somatic cells in mammals in which the cell has more than one X chromosome.4 In interphase, only one X chromosome usually functions in a cell at a given time; all other X chromosomes are present in the form of X-chromatin bodies.
The time of X inactivation is uncertain but is probably around the time of implantation. The choice of which X chromosome (i.e., maternal or paternal) is inactivated in a given cell is random if both X chromosomes are normal. After a given X is inactivated, however, the descendants of that cell retain the identical pattern. Because X inactivation occurs only after the embryo
contains hundreds of cells, normal females have two populations of cells: in one, the maternal X is active; in the other, the paternal X is active. The biochemical basis of X inactivation is not well understood, but the inactivated DNA appears to be more heavily methylated than in the active X. (See also ref. 4a.)
contains hundreds of cells, normal females have two populations of cells: in one, the maternal X is active; in the other, the paternal X is active. The biochemical basis of X inactivation is not well understood, but the inactivated DNA appears to be more heavily methylated than in the active X. (See also ref. 4a.)
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


