The anatomy and circulation of the spleen
The spleen lies under the left costal margin, has a normal weight of 150–250 g and a length of between 5 and 13 cm. It is normally not palpable but becomes palpable when the size is increased to over 14 cm.
Blood enters the spleen through the splenic artery which then divides into trabecular arteries which permeate the organ and give rise to central arterioles (Fig. 10.1). The majority of the arterioles end in cords which lack an endothelial lining and form an open blood system unique to the spleen with a loose reticular connective tissue network lined by fibroblasts and many macrophages. The blood re-enters the circulation by passing across the endothelium of venous sinuses. Blood then passes into the splenic vein and so back into the general circulation. The cords and sinuses form the red pulp which forms 75% of the spleen and has an essential role in monitoring the integrity of red blood cells (see below). A minority of the splenic vasculature is closed in which the arterial and venous systems are connected by capillaries with a continuous endothelial layer.
Figure 10.1 Schematic representation of the blood circulation in the spleen. Most blood flows in an ‘open’ circulation through splenic cords and regains entry into the circulation through the venous sinuses.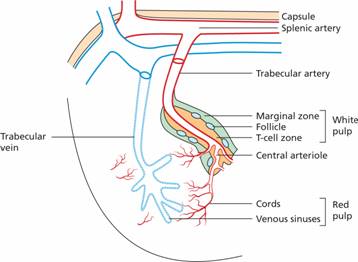
The central arterioles are surrounded by a core of lymphatic tissue known as white pulp which has an organization similar to lymph nodes (Fig. 10.1). The periarteriolar lymphatic sheath (PALS) lies directly around the arteriole and is equivalent to the T zone of the lymph node (see p. 128). B-cell follicles are found adjacent to the PALS and these are surrounded by the marginal zone and perifollicular zone which are rich in macrophages and dendritic cells. Lymphocytes migrate into white pulp from the sinuses of the red pulp or from vessels that end directly in the marginal and perifollicular zones.
There are both rapid (1–2 min) and slow (30–60 min) blood circulations through the spleen. The slow circulation becomes increasingly important in splenomegaly.
The spleen is the largest filter of the blood in the body and several of its functions are derived from this.
Control of red cell integrity
The spleen has an essential role in the ‘quality control’ of red cells. Excess DNA, nuclear remnants (Howell–Jolly bodies) and siderotic granules are removed (Fig. 10.2). In the relatively hypoxic environment of the red pulp, and because of plasma skimming in the cords, the membrane flexibility of aged and abnormal red cells is impaired and they are retained within the sinus where they are ingested by macrophages.
Figure 10.2 Splenic atrophy: peripheral blood film showing Howell–Jolly bodies, Pappenheimer bodies (siderotic granules; see p. 30) and misshapen cells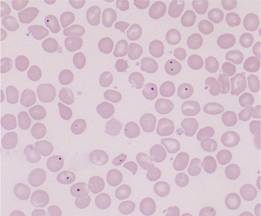
Immune function
The lymphoid tissue in the spleen is in a unique position to respond to antigens filtered from the blood and entering the white pulp. Macrophages and dendritic cells in the marginal zone initiate an immune response and then present antigen to B and T cells to start adaptive immune responses. This arrangement is highly efficient at initiating immune responses to encapsulated bacteria and explains the susceptibility of hyposplenic patients to these organisms.
The spleen, like the liver, undergoes a transient period of haemopoiesis at around 3–7 months of fetal life but is not a site of erythropoiesis in the adult. However, haemopoiesis may be re-established in both organs as extramedually haemopoiesis, in disorders such as primary myelofibrosis or in chronic severe haemolytic and megaloblastic anaemias. Extramedullary haemopoiesis may result either from reactivation of dormant stem cells within the spleen or homing of stem cells from the bone marrow to the spleen.
Ultrasound is the most frequently used technique to image the spleen (Fig. 10.3). This can also detect whether or not blood flow in the splenic, portal and hepatic veins is normal, as well as liver size and consistency. Computed tomography (CT) is preferable for detecting structural detail and any associated lymphadenopathy (e.g. for lymphoma staging). Magnetic resonance imaging (MRI) also gives improved fine detail structure. Positron emission tomography (PET) is used particularly for initial staging and for detecting residual disease after treatment of lymphoma (Fig. 10.4).
Figure 10.3 The spleen. (a) Ultrasound of spleen showing splenomegaly (15.3 cm). (b) Normal spleen (10 cm) on computed tomography (CT) scan. (c) CT scan: the spleen is enlarged and shows multiple low density areas. A diagnosis of diffuse large cell B lymphoma was made histologically after splenectomy. (Figs (a) and (b) courtesy of Dr T. Ogunremi.)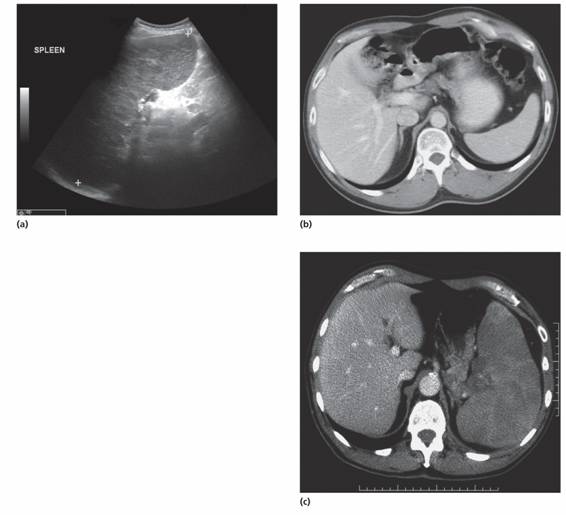
Figure 10.4 (a) Axial positron emission tomography (PET), (b) fused PET/CT and (c) CT images demonstrating a solitary focal area of [18
Related posts:
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


