Figure 38-1 Examples of Gleason grade 3, 4, and 5 prostate cancer. Gleason grade 3 shows well-formed, separate glands (A). Gleason grade 4 shows merging or cribriform glands (B). Gleason grade 5 is the most poorly differentiated, and cancer cells no longer form glands but are visible as sheets of cells (C).
Molecular Pathology
Hereditary Prostate Cancer
Family history is one of the strongest risk factors for the development of prostate cancer, with a two- to eightfold higher risk of prostate cancer in men with an affected first-degree relative. 6 Prostate cancer associated with familial clustering and high incidence of cancer among multiple first-degree relatives with a diagnosis before age 60 is considered hereditary and/or familial prostate cancer (HPC/FPC). Approximately 9% of all cases are attributable to hereditary prostate cancer following an autosomal dominant susceptibility pattern. Prostate cancer susceptibility genes have been identified using lineage analysis of affected families. Significant linkage between chromosome 1q24-25, the HPC1 locus, and hereditary prostate cancer has been established. RNASEL, which lies within the HPC1 locus, encodes an endoribonuclease that mediates the activities of an interferon-inducible RNA degradation pathway. Polymorphisms of the RNASEL gene have been associated with increased prostate cancer risk. However, not all studies have confirmed these findings. Mutations in the RNASEL gene do not occur at a greater frequency in patients with familial prostate cancer compared with patients with sporadic prostate cancer. Recent genome-wide association studies (GWAS) consistently identified that several single-nucleotide polymorphisms (SNPs) in the 8q24 locus are associated with risk of HPC/FPC22. Genetic variants caused by polymorphisms or mutations in other genes, such as PALB2, BRCA2, the androgen receptor (AR), 5-α-reductase type II (SRD5A2), and CYP17, have also been implicated in the development of HPC/FPC.
Table 38-1
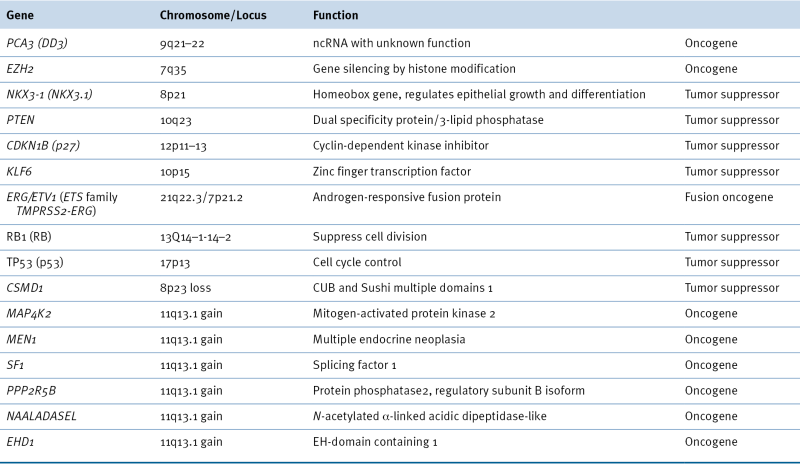
Oncogenes and Tumor Suppressors Implicated in Prostate Cancer
Gene Mutation
Early candidate gene approaches have implicated many different genes in prostate cancer. Germline mutations involving ELAC2 (HPC2), MSR1, and RNASEL genes have been reported in familial prostate cancer. The most common somatic mutations found in sporadic prostate cancer include TP53, PTEN, and AR. Recent whole-genome exon sequencing analyses identified significant mutated genes, including TP53, AR, ZFHX3, RB1, PTEN, APC, MLL2, OR5L1, and CDK12. Of these genes, MLL2, OR5L1, and CDK12 have unknown tumor suppressor functions in prostate cancer. 7 The most commonly affected signaling pathways by genetic alterations are the WNT signaling (TP53, APC, CTNNB1, MYC, and SMAD4) and the PTEN interaction network (PTEN, MAGI3, and HDAC11). 7
DNA Copy Number Variation
DNA copy number variation (CNV) is DNA structure alteration involving relatively large (at least 1 kb) regions. CNV can manifest as loss or gain of chromosomal regions. Earlier studies using fluorescence in situ hybridization (FISH) and comparative genomic hybridization (CGH) identified common losses at 1p, 6q, 8p, 10q, 13q, 16q, and 18q and gains at 1q, 2p, 7, 8q, 18q, and Xq. For example, MSR1, NKX3.1, and N33 are candidate tumor suppressor genes in prostate cancer lying within the most commonly deleted regions on chromosome 8p. MSR1 encodes a receptor on the macrophage cell surface that induces binding of oxidized low-density lipoprotein and other polyanionic ligands. Mutations, polymorphisms, or loss of the MSR1 gene may compromise global macrophage function, thereby exposing organs, including the prostate, to oxidative stress and damage. Although this gene does not code for prostatic proteins directly, oxidative stress has been implicated in the initiation of prostate carcinogenesis.
Loss of 8p23, a region that harbors the CUB and Sushi multiple domains 1 gene (CSMD1), has been associated with advanced prostate cancer. The retinoblastoma (RB1) gene is also a tumor suppressor gene and lies within the 13q locus. It is deleted in early prostate cancer development in animal models, prostate cancer cell lines, and some human prostate cancer specimens. RB1 inactivation in prostate cancer is the result of loss of heterozygosity and mutation. The 10q locus is lost in up to 45% of prostate cancers examined, and MXI1 and PTEN are two putative tumor suppressors in this region.
Regions of chromosome amplification in advanced prostate cancer include 8q containing the MYC gene and Xq11-13 encoding the AR gene. Gene amplification at 11q13.1 has been associated with disease recurrence. There are several candidate genes in this location (Table 38-1 ), but only MEN1 and MAP4K2 correlate with disease progression.
Recent prostate cancer genome sequencing studies consistently show that the genes most commonly affected by loss of copy number are CHD1, NTSE, PTEN, RB1, and TP53, and the most gained genes are AR, MYC, PIK3CA, and the HOXA3 cluster. These aberrations are more prominent in castration-resistant prostate cancer compared to localized disease. 7,8
Gene Fusion
The prostate cancer genome can harbor an average of 90 chromosomal rearrangements involving many genes. One of the most frequent gene fusion events in prostate cancer is the fusion of the TMPRSS2 and ERG genes, which are located 3 Mb apart on chromosome 21q22.2. 9 TMPRSS2 is an androgen-responsive gene, and ERG encodes an erythroblast transformation-specific (ETS) transcription factor. Their fusion is believed to be stimulated by androgens that recruit AR and TOP2B topoisomerase to chromosomal sites where TOP2B introduces double-strand breaks in DNA. TMPRSS2-ERG fusion results in overexpression of ETS genes in prostate cancer. TMPRSS2-ERG has been identified in 40% to 70% of prostate cancer and correlated with metastasis and disease-specific mortality. Gene fusion events such as TMPRSS2-ERG may be used for prostate cancer diagnosis through simple PCR detection of gene fusions in urine sediment.
WNT Signaling and β-Catenin
The Wnt signaling pathway plays a key role in embryonic development and is essential for the maintenance of stem cells. Wnt is an extracellular protein that interacts with the membrane-bound frizzled receptor to initiate its biologic activity. Wnt signaling leads to stabilization of CTNNB1 and its nuclear accumulation. Nuclear CTNNB1 converts the TCF/LEF DNA-binding protein complex from a transcriptional repressor into a transcriptional activator. Inappropriate activation of the Wnt pathway is observed in many cancers and is putatively associated with tumor development.
In mice, CTNNB1 stabilization through targeted excision of CTNNB1 exon 3 induces prostate intraepithelial neoplasia (PIN)-like lesions that are similar to the early stages of human prostate cancer. In human prostate cancers, high levels of nuclear CTNNB1 are detectable by immunohistochemistry, whereas their levels are undetectable in normal prostate tissue. High levels of CTNNB1 expression are associated with the more aggressive prostate tumors. Together, these findings imply that inappropriate activation of the Wnt signaling pathway can contribute to prostate cancer and progression.
There are several mechanisms by which the Wnt pathway may be inappropriately activated in prostate cancer; DNA methylation plays a key role in several of these processes. The adenomatous polyposis coli (APC) gene is hypermethylated in prostate tumors relative to samples of benign prostatic hyperplasia (BPH; 64.1% vs. 8.7%). APC is a key component of the CTNNB1 degradation complex. Thus, methylation-dependent silencing of APC can lead to CTNNB1 accumulation and Wnt pathway activation.
E-cadherin (CDH1), a cell membrane protein, interacts with CTNNB1 and sequesters it at the inside surface of the cellular membrane. However, CDH1 expression is often lost in prostate cancers because of chromosomal loss or promoter hypermethylation. Thus, because CDH1 is no longer present, CTNNB1 is released into the cytoplasmic and nuclear compartments, leading to Wnt pathway activation. Finally, the secreted-frizzled related proteins (SFRPs) and Wnt inhibitory factor-1 (Wif-1) sequester Wnt and antagonize Wnt signaling. In this manner, loss of SFRP/Wif-1 expression can lead to Wnt pathway activation. The genes encoding several of the SFRPs and Wif-1 are epigenetically silenced by DNA methylation in colorectal, lung, bladder, and kidney cancers and lymphocytic leukemia. In prostate cancer, Wif-1 expression is strongly suppressed. The SFRP1 gene is also aberrantly hypermethylated in prostate tumors relative to BPH tissue and is partially to completely methylated in several human prostate cancer cell lines. These findings suggest that silencing of genes antagonist to Wnt may play a role in prostate cancer development.
The Wnt signaling pathway may interact with other signaling pathways such as the AR-CTNNB1, which in turn can upregulate AR transcriptional activity in an androgen-dependent manner. Subsequently, AR enhances nuclear translocation of CTNNB1. In addition, PI3K/Akt can modulate the activity of CTNNB1 by phosphorylation of CTNNB1 by GSK3B, a substrate of Akt.
MicroRNA and Other Noncoding RNA in Prostate Cancer
Only a small fraction of the transcription output in human genome encodes for proteins. Noncoding RNAs (ncRNAs) are arbitrarily classified into two major classes based on their size: small (microRNA) and long ncRNA (lncRNA).
MicroRNA and Prostate Cancer
Small ncRNAs, exemplified by microRNA (miRNA), are known to regulate diverse biological processes in stem cells, development, differentiation, metabolism, and disease such as cancer. Several hundred miRNAs have been identified in human cells, and these are transcribed from the genome as long, primary miRNAs ranging in size from hundreds to thousands of nucleotides. miRNAs depend on multiple proteins for their biogenesis and function. Aberrations in any of these proteins will affect miRNA-mediated gene regulation. Expression of Dicer, a key gene in the biosynthesis of miRNA, is upregulated in a significant fraction of prostate cancer and is associated with aggressive cancer features. Knockout of Dicer in the mouse prostate impairs prostate stem cell activity and causes prostate atrophy. Some other components of the miRNA machinery (XPO5, Ago1, Ago2, HSPCA, MOV10, and TNRC6B) are also upregulated in prostate cancer.
Processed miRNAs have been found to regulate gene expression by inhibiting protein translation and/or enhancing the degradation of target gene mRNA with which the miRNAs have imperfect sequence complementarity in the 3′UTR region. It has been predicted that each miRNA can target hundreds of genes, and a third of human protein-coding genes are regulated by miRNA. An increasing body of evidence suggests that miRNAs are involved in the initiation and development of different types of cancers, including prostate cancer.
Expression Signature of miRNA in Prostate Cancer
High-throughput miRNA expression profiling studies have found altered miRNA expression in prostate cancer, providing evidence for the involvement of miRNAs in this disease. miRNAs that have increased expression or amplification in cancer can act as oncogenes to enhance cell proliferation and survival by the inhibition of protein-coding tumor suppressor genes. They are thus called oncomiRs. Several such oncomiRs have been identified in prostate cancer cell lines including miR-21, miR-291, miR-221, miR-222, and the well-known miR-17-92 cluster, which contains six members: miR-17, miR-18a, miR-19a, miR-20a, miR-19b-1, and miR-92a-1. Some common target genes of these oncomiRs include PTEN, BIM, RB1, p21, and p27. On the other hand, miRNAs that normally target and suppress are regarded as tumor suppressor miRNAs (ts-miRNAs). In prostate cancer, the miR-15a/16-1 cluster that targets BCL2 and CCND1 is downregulated in prostate cancer. Forced expression of these miRNAs can lead to suppression of cell proliferation and induction of apoptosis. 10 Other commonly identified tumor suppressor miRNAs include miR-34a/b/c, miR-145, miR-205, and let-7.
miRNA and the AR Signaling Pathway
The AR signaling pathway and miRNAs are engaged in reciprocal regulation via multiple interaction points. miRNA can suppress AR expression through the canonical miRNA pathway or affect AR transcriptional activity by modulating other AR cofactors such as MYC. A systemic analysis of miRNAs identified a number of miRNAs that could suppress AR expression post-transcriptionally by targeting the AR 3′UTR element. Similar to protein coding genes, miRNA genes are also under the regulation of many transcription factors, including AR, which binds to ARE in the promoter of many miRNAs. As the first example, AR can bind to the ARE of miR-21, a prostate oncomiR, and directly regulate its expression as a downstream signaling effector. 11 Several other miRNAs were later found to be also under the regulation of AR, including miR-101, miR-141, miR-27a, miR-32, and miR-148a.
miRNA as Biomarkers for Prostate Cancer Diagnosis
miRNAs derived from cancer cells can exist as circulating extracellular or cell-free RNA and are remarkably stable in body fluids such as plasma/serum. These extracellular miRNAs can be detected and quantified using either PCR-based methods or other high-throughput approaches, including miRNA microarray and next-generation sequencing. A number of studies have performed miRNA profiling in body fluids including serum, plasma, and urine and have identified several miRNAs whose levels can serve as diagnostic and/or prognostic markers for prostate cancer patients. 12,13 Some of the miRNAs that consistently exist in prostate cancer patients across different studies include miR-141 and miR-375.
lncRNA and Prostate Cancer
The longer class of ncRNAs is known as long intergenic RNA (lncRNA), with sizes ranging from a few hundred to thousands of nucleotides. lncRNAs can regulate protein-encoding genes by affecting transcription and chromatin state by mechanisms distinct from those used by small ncRNAs. Thus genetic alterations and aberrant expression of lncRNAs can be a causal factor in disease. Several lncRNAs pertinent to prostate cancer have been identified. A 3.7-kb lncRNA known as PCA3 (DD3) has been mapped to chromosome 9q21-22 and shown to be highly overexpressed in prostate cancer samples. Genetic variation in lncRNAs has been found to affect prostate cancer risk. Microarray profiling of intronic transcripts identified many ncRNAs expressed in prostate cancer samples, and the expression levels of some correlate with the extent of prostate tumor cell differentiation. 14 By high-throughput RNA sequencing of 102 prostate tissues and cell lines, Prensner and colleagues identified 121 unannotated prostate cancer–associated ncRNA transcripts (PCATs) and have characterized one of them, PCAT-1, as a prostate cancer–specific ncRNA functionally implicated in disease progression.
Given the regulatory role of lncRNA in prostate cancer, their aberrant expression suggests that they may serve as tumor biomarkers. In this regard, Cui and co-workers identified an lncRNA termed PlncRNA that expresses higher in prostate cancer cells compared to normal prostate epithelial cells. 15 Knockdown of PlncRNA has an inhibitory effect on prostate cancer cells, potentially through its regulation of AR activity
Epigenetic Alterations in Prostate Cancer
Epigenetic alterations contribute to the malignant transformation and progression of prostate cancer. Figure 38-2 shows the potential contribution of epigenetic events to the development of prostate cancer and its progression to advanced and castration-resistant cancer. Initially DNA methylation was regarded as a new type of promising biomarker for prostate cancer diagnosis and prognosis, and a therapeutic target. Despite intensive research efforts in the past decade, results from clinical studies evaluating DNA methylation as a biomarker have been disappointing, and so far no biomarker has advanced into the clinical arena.
Hypermethylation
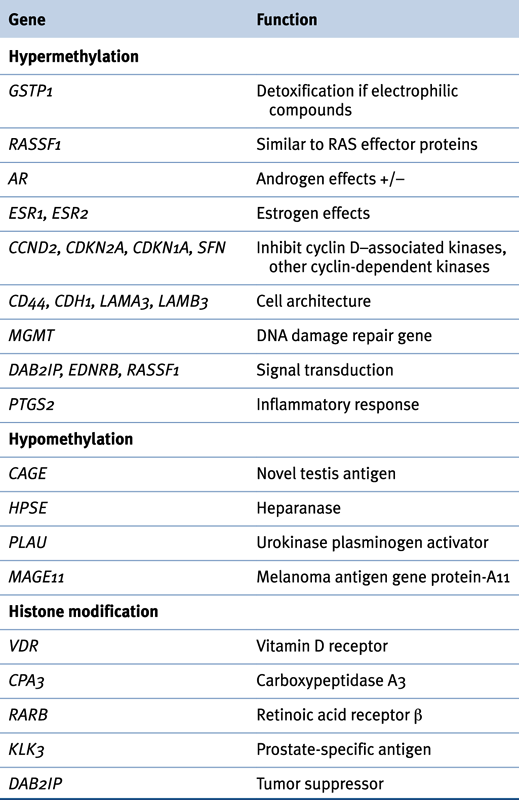
DNA hypermethylation is one of the most common and best-characterized epigenetic abnormalities in prostate cancer. Genes including classic and putative tumor suppressor genes as well as genes involved in a number of cellular pathways, such as hormonal responses, cancer cell invasion/tumor architecture, cell cycle control, and DNA damage repair, can be hypermethylated in prostate cancer. For many of these genes, promoter hypermethylation is often the mechanism responsible for their functional loss in prostate cancer. Inappropriate silencing of these genes can contribute to cancer initiation, progression, invasion, and metastasis. Some commonly hypermethylated genes in prostate cancer are discussed next (Table 38-2 ).
DNA Damage-Repair Genes
Antioxidants and DNA repair pathways protect the genome and maintain genome stability during replication or following DNA damage. Hypermethylation of genes important for such processes, such as glutathione S-transferase Pi (GSTP1) and O-6-methylguanine DNA methyltransferase (MGMT), has been frequently documented in prostate cancer. GSTP1, located at chromosome 11q13, belongs to a supergene family of glutathione S-transferases (GSTs) that play an important role in the detoxification of carcinogens and cytotoxic drugs by catalyzing their conjugation to glutathione. GSTP1 inactivation may lead to increased cell vulnerability to oxidative DNA damage and the accumulation of DNA base adducts, which can precede carcinogenesis.
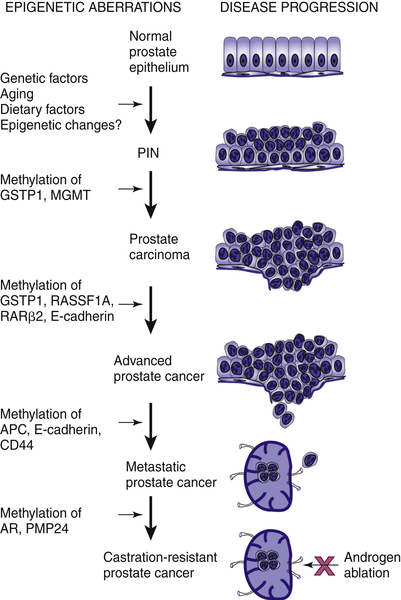
Figure 38-2 Epigenetic alterations in the development of prostate cancer Multiple factors are associated with the development of prostate cancer, including genetic predisposition, environmental factors, diet, ethnicity, and aging. Many of these factors modify the genome through epigenetic effects, and DNA methylation may be an early event causing inactivation of DNA damage repair genes such as GSTP1 and MGMT. Subsequent inactivation of cell-cycle control genes provides a growth advantage leading to locally advanced prostate cancer. Functional loss of genes in the cell adhesion pathway, such as CD44, may allow for metastasis. Ultimately, inactivation of AR via DNA hypermethylation allows cancer cells to become androgen insensitive. From Li LC, Okino ST, Dahiya R. DNA methylation in prostate cancer. Biochim Biophys Acta. 2004;1704:87-102, with permission.
In prostate cancer, methylation of the GSTP1 gene promoter is the most frequently detected epigenetic alteration. Elevated CpG methylation has been detected in prostate cancer tissues as early as at the stage of atypia and PIN. GSTP1 promoter hypermethylation can also be readily detected in the serum/plasma, urine, and ejaculates of prostate cancer patients with high specificity but unsatisfying sensitivity. Current findings from a large number of studies do not support the feasibility of using GSTP1 as an independent tumor biomarker, but it may complement prostate-specific antigen (PSA) screening for prostate cancer diagnosis.
Table 38-2
Epigenetic Changes in Prostate Cancer
Hormone Receptors
AR mediates testosterone and dihydrotestosterone activity, which is essential for the development and maturation of the prostate gland and prostate cancer. Most prostate cancer is initially androgen dependent, but eventually becomes androgen independent after androgen-deprivation therapy. Androgen-independent prostate cancers are characterized by a heterogeneous loss of AR expression. Genetic alterations that alter the sensitivity of the receptor to androgen, such as AR gene mutation and, more commonly, amplification without loss of AR expression, are thought to play key roles in the development of androgen-independent prostate cancer. The prostate expresses two types of estrogen receptors (ERs): ERα (ESR1) and ERβ (ESR2), whose functional role in the prostate and prostate cancer remains controversial. Hypermethylation of ERs leading to decreased expression increases with aging and can contribute to tumor progression in prostate cancer patients.
Cell Cycle Control Genes
An important characteristic of tumor cells is unbalanced proliferation due to impaired regulation of the cell cycle. The multiple checkpoints that control the cell cycle include the retinoblastoma protein, cyclins, cyclin-dependent kinases (CDKs), and CDK inhibitors (CDKIs). CDKIs are potential tumor suppressor genes that act as molecular brakes on cell cycle progression. Failure of cell cycle arrest due to alterations in CDKI expression has been implicated in prostate cancer. CDKIs are grouped into two families: the INK4 family and the CIP/KIP (kinase inhibitor protein) family. The INK4 family includes CDKN2A (p16), CDKN2B (p15), CDKN2C (p18), and CDKN2D (p19) and inhibits cyclin D–associated kinases (CDK4 and CDK6). The CIP/KIP family, which includes CDKNIA (p21), CDKN1B (p27), and CDKN1C (p57), inhibits most CDKs.
CDKN2A can be inactivated in prostate cancer by a variety of mechanisms, including deletion, point mutation, and hypermethylation. Methylation-mediated inactivation of the CDKN2A gene has been reported in prostate cancer cell lines and tissues at a very low frequency (0% to 16%). Inactivation of other cell cycle genes such as CDKN2B, CDKN1A, and CDKN1B by hypermethylation is rare in prostate cancer.
The Ras-association domain family-1 gene (RASSF1) is located at 3p21.3 and encodes a protein similar to the RAS effector proteins. A tumor suppressor role has been proposed for RASSF1. RASSF1 promoter methylation is a common event in prostate cancer and high-grade PIN, occurring in 54% to 96% of tumors, and increases with higher Gleason scores.
Tumor Invasion and Tumor Architecture Genes
The cadherin-catenin adhesion system is critical for the preservation of normal tissue architecture and is regulated by a family of proteins collectively termed cell adhesion molecules (CAMs). Decreased expression of CDH1 and other CAMs has been reported to have prognostic significance in various human cancers, including prostate cancer. In prostate cancer, expression of CDH1 is markedly suppressed and its promoter is methylated to varying degrees. In addition, methylation of the CDH1 promoter is increased in advanced prostate cancer, making it a potential biomarker for cancer progression.
CD44 is an integral membrane protein involved in matrix adhesion and signal transduction. Loss of CD44 expression correlates with methylation of its gene promoter in prostate cancer and is associated with stage and prognosis. Other genes involved in the cadherin-catenin adhesion system have also shown methylation-mediated inactivation in prostate cancer, such as H-cadherin, adenomatous polyposis coli (APC), caveolin-1 (CAV1), laminin α-3 (LAMA3), laminin β-3 (LAMB3), and laminin γ-2 (LAMC2).
Other Putative Tumor Suppressor Genes
Other possible tumor suppressor genes that are subject to epigenetic inactivation in prostate cancer include KAI1 (a prostate-specific tumor metastasis suppressor gene), inhibin-α (a member of the TGF-β family of growth and differentiation factors), and DAB2IP, a novel GTPase-activating protein for modulating the Ras-mediated signal pathway. 16 Recent high-throughput DNA methylation profiling in prostate cancer cell lines and tissues unveiled methylated genes such as SLC15A3, KRT7, TACSTD2, GADD45b 17 OXD3 and BMP7 18 PR83, ADCY4, LOC63928, and D4S234E; 19 however, these genome-wide analyses seem not to have yielded consistent information identifying new prostate cancer biomarkers.
Related posts:



Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


