Fig. 8.1
The luteostatic “endocrine–exocrine” model for maternal recognition of pregnancy in the pig. In cyclic gilts, the loss of PGR in the uterine epithelia by day 10 of the estrous cycle is temporally associated with increased expression of PTGS1 and PTGS2 by uterine LE and GE which results in the release of PGF2α and some PGE. In the absence of conceptuses or estrogen stimulation, the majority of the PGF2α produced is released into the uterine capillary bed (endocrine direction) and then into the uterine vein and general circulation to exert luteolytic effect on both ovaries. The systemic effect of PGF2α results as only 30 % of PGF2α is metabolized in one passage through the lungs. In pregnant gilts, conceptuses release estrogens stimulating secretion of the majority of the endometrial PGF2α into the uterine lumen where it is metabolized by the conceptus trophoblast to inactive form or PGE2. In addition, IL1B2 produced by the conceptuses may increase expression of endometrial PGE synthase-1 to increase the ratio of PGE2 to PGF2α and sequester prostaglandins in the uterine lumen or the endocrine release of PGE2 may support CL function during early pregnancy
In addition to the alteration of PG movement from the uterine endometrium, release of estrogen from the expanding conceptuses can have a direct luteotrophic effect on the CL (Conley and Ford 1989) as well as stimulating a localized increase in endometrial vascular permeability (Keys and King 1988, 1995) and an overall increase in uterine blood flow (Ford et al. 1982; Ford and Stice 1985). During conceptus elongation and the early peri-implantation period, the endometrium increases the release of a number of growth factors and cytokines such as epidermal growth factor (EGF), insulin-like growth factor-1 (IGF-1), fibroblast growth factor 7 (FGF7), vascular endothelial growth factor (VEGF), interleukin 6 (IL-6), transforming growth factor beta (TGFβ), and leukemia inhibitory factor (LIF) (see review Bazer et al. 2010; Geisert et al. 2014). Endometrial release of EGF, FGF7, LIF, and IGF-1 is specifically enhanced in the epithelium during the period of conceptus elongation and estrogen secretion. Conceptus estrogens also increase endometrial expression of genes for aldo–keto reductase (Seo et al. 2014a), interleukin 1 receptor accessory protein (IL1RAP) (Seo et al. 2012a), lysophosphatidic acid receptor 3 (LPAR3) (Seo et al. 2008), secreted phosphoprotein 1 (SPP1) (White et al. 2005), and stanniocalcin1 (STC1) (Song et al. 2009).
8.3.3 Rapid Elongation of the Conceptus
The capacity of pig conceptuses to secrete estrogen clearly provides the major mechanism by which the movement of PGF2α is altered from endocrine to exocrine by the uterine endometrium. However, in order for conceptus estrogens to reduce PGF2α release into the utero-ovarian vein, the conceptuses must first cover the majority of the uterine surface area after day 11 of pregnancy (Dhindsa and Dziuk 1968b). Heuser and Streeter (1929) were the first to document the spherical, tubular, and filamentous morphological forms of the pig conceptus. In addition to pig conceptuses expanding across the uterine surface to inhibit luteolysis, trophoblast expansion provides sufficient surface area for nutrient transfer through the trophectoderm initially and then the diffuse, epitheliochorial placenta.
The peri-implantation pig conceptus undergoes a remarkable transformation from a 1–2 mm sphere to a 9–10 mm long ovoid shape between days 10 and 12 of pregnancy and then rapidly remodels to tubular and filamentous forms by elongating at 30–40 mm/h to >100 mm in length in 1–2 h (Anderson 1978; Geisert et al. 1982b). Perry (1981) stated that “the rapidity of elongation and reduction in diameter that accompanies it suggest that it is by deformation rather by cell division.” The rapid alteration in conceptus morphology within the uterine horns does occur through cellular migration and remodeling of the trophectoderm and endoderm following differentiation of the epiblast mesoderm (Geisert et al. 1982b; Mattson et al. 1990). Many of the morphological forms of the conceptus can be found within the same uterus (Anderson 1978) indicating that development of pig conceptuses within a litter is not necessarily uniform and that may contribute to some of the early embryonic losses that occur in pigs (Pope 1994). Upon completion of rapid elongation, the filamentous conceptuses continue to elongate to 60 cm by day 13 and reach a length of 1 m by day 18 of gestation. Because the placental membranes of pigs do not overlap, females with a high ovulation rate have conceptuses that must compete for adequate uterine space for development and survival to term. The variation in conceptus development on day 12 may provide some of the selection pressure for survival of conceptuses that are first to elongate and establish their surface area for implantation and placentation in the uterus (Pope 1994).
Although growth of the pig conceptuses is regulated through release of uterine growth factors during early development (Geisert et al. 2014), rapid trophoblast elongation and estrogen secretion are triggered by conceptus development and cellular differentiation (Fig. 8.2). Mesoderm differentiation and outgrowth from the epiblast are markers for the increase in steroidogenesis of the developing spherical conceptuses (Yelich et al. 1997; Conley et al. 1992, 1994). The growth and expansion of mesoderm between the trophectoderm and underlying extraembryonic endoderm may provide the cellular interactions needed to initiate the elongation process (Fig. 8.2). Cellular alterations in junctional complexes of the trophectoderm and migration of endodermal cells involved in conceptus elongation occur at the epiblast and extend down the “elongation zone” to the tips of the tubular conceptus (Geisert et al. 1982b). Epiblast production of FGF4 and the activation of mitogen-activated protein kinases (MAPK) through trophectoderm expression of fibroblast growth factor 2 (FGFR2) could induce expression of bone morphogenetic protein 4 (BMP4) by the mesoderm (Valdez Magaña et al. 2014). It is quite possible that paracrine secretion of BMP4 from the developing mesoderm initiates pathways to induce the cellular changes required for localized migration of the underlying endoderm and modification of microfilaments and junctional complexes (Mattson et al. 1990) making the overlying trophectoderm layer more fluid for rapid remodeling during elongation. Certainly, Perry’s (1981) suggestion that “conceptus elongation occurs in much the same way as does a ball of plasticene rolled under the hand” fits with the centralized (epiblast) localization of cell migration and shifting of cells needed for rapid transformation of ovoid to filamentous forms of the conceptuses (Fig. 8.2).
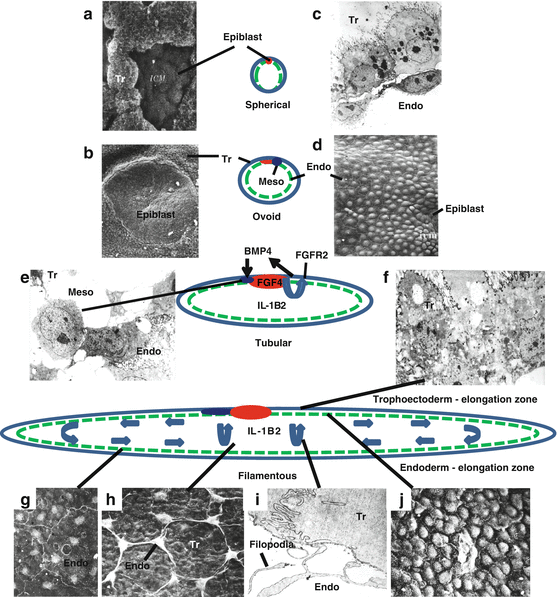
Fig. 8.2
Elongation of the trophoblast of pig conceptuses involves cellular differentiation and rapid remodeling of the trophectoderm (Tr) and endoderm (Endo) between days 11 and 12 of pregnancy. The trophectoderm covering (Rauber’s layer) the early spherical (2–4 mm) conceptus is removed (a) exposing the epiblast (ICM) on the surface by the 5 mm spherical stage (b). The early spherical conceptus is composed of outer rounded cuboidal trophectoderm with a closely attached layer of endoderm on its basement membrane (c). The endoderm facing the inner blastocele of the conceptus forms a continuous layer from the epiblast (d). Differentiation of conceptus mesoderm (Meso) is evident after the 5 mm spherical conceptus stage when estrogen and IL1B2 production are first detected (f). Epiblast production of FGF4 is proposed to stimulate fibroblast growth factor receptor 2 (FGFR2) receptors present in the trophectoderm to stimulate BMP4 production required for differentiation of mesoderm cells. Increased expression of IL1B2 mRNA and protein by tubular conceptuses is proposed to trigger cellular movement and remodeling during elongation of the pig conceptus. Near the epiblast, cellular junctional complexes of the trophectoderm undergo a shift to allow cell movement and transition of those cells to a columnar shape (e) and the underlying endoderm cells form filopodia (h, i) that pull the overlying trophectoderm toward the elongation zone. Movement of the spindle-shaped endodermal cells toward the elongation zone tightly compacts the endoderm cells together (j). With the alteration in shape of trophectoderm and increased density of the endoderm in the elongation, the cells are moved toward the tips of the elongating conceptus returning the cells to normal pattern (g) as the process repeated until elongation is complete
8.3.4 Expression of Interleukin 1 Beta (IL1B) by Conceptuses
Analyses of the transcriptome of developing spherical, ovoid, tubular, and filamentous pig conceptuses have provided information concerning genes involved during this critical period in development (Ross et al. 2003a, b, 2009; Blomberg et al. 2005, 2006). Although transcriptome profiling identified a number of genes that are up- or downregulated during this critical period of conceptus transformation, IL1B is the most abundantly expressed transcript during the time of tubular to filamentous transition for pig conceptuses (Ross et al. 2009). Expression of IL1B during the period when pig conceptuses transition to filamentous morphology was first described by Tuo et al. (1996). Because the rapid increase in IL1B mRNA and IL1B protein expression during conceptus elongation is immediately followed by a loss of transcript expression, IL1B was proposed as a possible candidate for initiating the cellular signaling pathway for conceptus remodeling (Fig. 8.2). Interestingly, the pig conceptus expresses a novel isoform, IL1B2, which resulted from gene duplication (Mathew et al. 2015). Expression of conceptus IL1B2 is specific to the pig conceptus as the transcript is not detected in peripheral tissues or in other mammals. Protein sequences of IL1B and IL1B2 are 85 % identical. Substitutions in amino acids may affect caspase-1 cleavage sites needed for IL1B2 secretion and biological activity. In addition, differences in the promoter region may contribute to specific expression of IL1B2 in the pig conceptus. IL1B promotes motility during human cytotrophoblast formation and induces secretion of urokinase plasminogen activator involved with endometrial invasion (Prutsch et al. 2012). Conceptus production of plasminogen activator is associated with the period of conceptus elongation, estrogen synthesis, and IL1B2 release in pigs (Fazleabas et al. 1983). IL1B can also stimulate phospholipase A2 (PLA2) (Kol et al. 2002) which increases membrane fluidity through the release of arachidonic acid from the phospholipid bilayer of the cell membrane. The increase of plasminogen activator and PLA2 activity during elongation of the conceptus (Davis et al. 1983) would be consistent with a role for IL1B2 in the induction of rapid cellular remodeling. The release of arachidonic acid from the conceptus trophoblast contributes to increases in secretion of PGs from the conceptus during and following elongation as conceptus expression of PTGS2 also increases (Wilson et al. 2002). However, PLA2 release of arachidonic acid appears to be the critical event in promoting membrane fluidity, as inhibition of PTGS1 and PTGS2 (downstream of PLA2) does not block conceptus elongation (Geisert et al. 1986).
8.3.5 Endometrial Stimulation by IL1B2
Although the dramatic increase in IL1B2 mRNA expression by pig conceptuses only occurs during the period of rapid elongation (day 11–12), IL1B2 protein is detectable in the uterine lumen from day 12 to 18 of gestation (Ross et al. 2009). Salivary lipocalin (SAL1), which functions as a transporter of hydrophobic compounds in biological fluids, is induced by IL1B and secreted by the uterine glands during conceptus elongation in the pig (Seo et al. 2011). Conceptus-induced secretion of SAL may play an important role in binding to lipids, PGs, and lysophosphatidic acids for implantation, placentation, and establishment of pregnancy in the pig (Seo et al. 2008).
Pro-inflammatory effects of the IL1family of cytokines on tissue are well established (Dinarello 2009). Tissue responses to IL1B are regulated through two receptors (IL1R1 functional, IL1R2 pseudo-receptor), receptor accessory protein and receptor antagonists (Dinarello 2009). Binding of IL1B2 to IL1R1 on the epithelial surface initiates a cascade of signaling pathways through activation of the inducible transcription factor, nuclear factor kappa-B (NFKB) (Hayden and Ghosh 2012). NFKB plays an essential role in sensing and adapting to alterations in the microenvironment of the immune system at the level of tissues and epithelia (Wullaert et al. 2011). Inactive dimers of NFKB are sequestered in the cytoplasm until receptor binding triggers phosphorylation and release of IKB (a regulatory protein inhibitor of κB) allowing NFKB translocation to the nucleus to effect gene transcription (Dinarello 2009).
Downregulation of PGRA in the endometrial LE and GE during the peri-implantation period not only plays a role in the loss of mucin-1 expression to open the window for conceptus attachment to uterine LE but also removes PGR-dependent inhibition of NFKB activation (Ross et al. 2010; Mathew et al. 2011). There is a clear spatiotemporal relationship between PGR downregulation and NFKB activation by IL1B2 from pig conceptuses (Mathew et al. 2011). Conceptus elongation and IL1B2 release stimulate nuclear translocation of NFKB in LE adjacent to conceptuses during the establishment of pregnancy (see review Geisert et al. 2014) Nuclear of translocation of the NFKB dimers stimulates transcription pathways involved in inflammation, cell adhesion, cytokine release, anti-apoptotic factors, and immunoreceptors (Hayden and Ghosh 2012). Many cytokines (TNFα, IL1, IL2, IL6, IL12, LIF, and GMCSF), chemokines (IL8 and RANTES), and PTGS2 are transcriptionally regulated by NFKB (Ali and Mann 2004). Activation of inflammatory pathways in the endometrium must be tightly regulated in order to prevent severe inflammation leading immunological rejection of the conceptus. Estrogen has the capacity to act as either an antagonist (Quaedackers et al. 2007) or agonist of NFKB activity (King et al. 2010). The presence of ESR1 in the endometrial LE and GE and tight coupling of conceptus estrogen synthesis and release with expression of IL1B2 would modulate the pro-inflammatory reaction of the uterus during elongation and placental attachment (see review Geisert et al. 2014).
Production of estrogen and IL1B2 by pig conceptuses enhances endometrial production of LIF and PTGS2. Endometrial LIF is associated with implantation in a number of species and increases on day 12 of pregnancy in the pig (see review Geisert et al. 2014). Inhibition of PG synthesis during the pre-implantation period causes early embryonic loss (Kraeling et al. 1985). IL1B stimulates endometrial PG synthesis and release, which, in addition to conceptus estrogens, may induce the pregnancy-specific increase in endometrial and uterine luminal content of PGE2 and PGF2α (Franczak et al. 2010; Seo et al. 2012a, 2014a). Increases in PGE2 production may occur through stimulation of endometrial PGE synthase-1(PGES-1) by conceptus IL1B2 (Franczak et al. 2010). An increase in the ratio of conceptus and endometrial PGE2 to PGF2α has been proposed to function as part of the luteostatic mechanism to sequester and metabolize PGF2α in the uterine lumen (Waclawik et al. 2009). In addition, an increase in PGE release into the utero-ovarian vein (Fig. 8.1) may have a direct luteotrophic action on the CL (Christenson et al. 1994). The PG transporters, ATP-binding cassette, subfamily C, member 4 (ABCCA4), and solute carrier organic anion transporter family, member 2A1 (SLCO2A1) are induced by IL1B and expressed in a temporal- and cell-specific manner within the endometrium of pregnant pigs (Seo et al. 2014b). Expression and cellular localization of the PG transporters within the endometrial LE and GE add support to the endocrine–exocrine model for the establishment and maintenance of pregnancy.
8.3.6 Secretion of Interferons by Conceptuses
Immediately following conceptus elongation and the rapid decline in IL1B2 gene expression, pig conceptuses express and secrete interferon gamma (INFG) and delta (INFD) between days 12 and 20 of gestation when there is a 567-fold increase in IFNG mRNA during the transition from spherical to day 14 filamentous conceptuses (La Bonnardière et al. 1991; Cencic and La Bonnardière 2002; Ross et al. 2009). Indeed, pig conceptuses are unique in secreting both type I and type II IFNs during the peri-implantation period of pregnancy. In contrast to sheep conceptuses which secrete type I IFN tau (IFNT), the pregnancy recognition signal in ruminants (Spencer et al. 2007), IFNs produced by pig conceptuses do not appear to be antiluteolytic (Harney and Bazer 1989; Lefèvre et al. 1998). Although pig conceptus IFNs are not known to influence pregnancy recognition, paracrine and autocrine effects of IFND and IFNG are suggested by localization of both the type I IFN receptor1 (IFNAR1, which binds pig IFND) and the type II IFN gamma receptor 1 (IFNGR1) on endometrial epithelial and conceptus trophectoderm (Niu et al. 1995; Lefèvre et al. 1998; D’andrea and La Bonnardiere 1998). When conceptus secretory proteins containing IFND and IFNG were infused into uteri of pseudopregnant pigs, uterine secretion of prostaglandin E2 was increased (Harney and Bazer 1989), as was expression of several IFN-responsive genes in the endometrium, including but not limited to signal transducer and activator of transcription 1 (STAT1) and STAT2, interferon regulatory factor 1 (IRF1), swine leukocyte antigens 1, 2, 3, 6, 7, and 8 (SLAs 1, 2, 3, 6, 7, 8), and beta 2 microglobulin (B2M) (Joyce et al. 2007a; Joyce et al. 2007b; Joyce et al. 2008; Johnson et al. 2009). Ka et al. (2009) reported that SLA-DQA, a major histocompatibility complex (MHC) class II gene, is expressed in the uterine endometrium at the time of conceptus implantation in pigs, and using endometrial explant cultures from day 12 of the estrous cycle, determined that expression of SLA-DQA and SLA-DQB mRNAs increased in response to IFNG (Kim et al. 2012).
Secretion of estrogens, IFND, and INFG by pig conceptuses could coordinate STAT1 activation in uterine LE and stroma to induce cellular pathways for trophoblast attachment and regulation of the maternal immune response to the semi-allogenic conceptuses (Joyce et al. 2007a, b). Interestingly, with the decline in IL1B2 secretion, there is an increase in endometrial IL18 expression during the period of trophoblast attachment from day 13 to 18 of gestation (Ashworth et al. 2010b). Previously referred to as interferon inducing factor, IL18 is a member of the pro-inflammatory IL1B family. A pregnancy-specific increase in endometrial caspase-1 expression increases release of IL18 into the uterine lumen which could stimulate INFG and IFND production by conceptuses. The switch from conceptus IL1B2 to endometrial IL18 production parallels the second sustained increase in conceptus estrogen production which suggests that estrogen modulates the endometrial response to the pro-inflammatory cytokines.
Fragmentation and loss of conceptuses following premature exposure of pregnant gilts to estrogen (day 9) are associated with lack of expression of IFNG by conceptuses required to activate STAT1 in stromal cells (Joyce et al. 2007b). The failure of IL18 to accumulate in the uterine lumen of estrogen-treated gilts is consistent with its role in stimulating IFNG production by pig conceptuses (Ashworth et al. 2010b). These studies illustrate the spatiotemporal sensitivity of the endometrial/conceptus interface to the interplay between estrogen and the expression of endometrial IL18 and IFNG by conceptuses during the period of implantation/placentation and maintenance of pregnancy in the pig.
8.4 Adhesion Cascade for Implantation
8.4.1 Implantation
Implantation of the conceptus and development of a placental connection to the maternal circulation are strong evolutionary advantages of eutherian mammals. The placenta (chorion, allantois, and amnion) forms the interface between the microcirculatory systems of the mother and conceptus and functions for efficient, sustained, and high-throughput exchange of nutrients, respiratory gases, and metabolic wastes, and it protects the growing embryo/fetus and is a source of hormones. Due to its recent appearance in the evolutionary record, a considerable variability exists among species relative to histogenesis and organization of the placenta (Carter and Enders 2013). The placental membranes are formed from components of the blastocyst and embryo; however, both the embryo and maternal endometrium begin to form components of the placenta as soon as the conceptus trophectoderm attaches to the endometrium. Indeed, the placental trophoblast interaction with maternal tissues remains extensive in all species. Blastocysts of some species, including primates and rodents, are invasive and penetrate the epithelial layer of the endometrium. Pigs, however, employ a vastly different strategy, and demonstrate a true epitheliochorial placentation in which there is no displacement or invasion of the maternal epithelium and the conceptus remains within the uterine lumen throughout gestation (Burton 1992). The changes that occur at the interface between trophectoderm and uterine LE during the initial stages of epitheliochorial placentation in pigs, day 13–26, have been elegantly described by Dantzer (1985). Throughout implantation, the glycocaylx that extends from the apical surface of the uterine LE is thicker than the glycocaylx at the surface of conceptus trophectoderm. On days 13 and 14, the uterine LE develops protrusions that become enclosed by caps of trophectoderm cells that serve to physically immobilize the conceptus; and by day 14, there is close apposition between the apical plasma membranes of trophectoderm and uterine LE cells. Interdigitating microvilli form between these plasma membranes through days 15 and 16, and then, the interface becomes increasingly complex as it functionally transitions from histotrophic to histotrophic and hemotrophic nutrient transport over day 15–20. This transition is characterized by the development of apical domes on the uterine LE that are closely related to the trophectoderm and provide long cytoplasmic extensions into a luminal space between the apical domes. Finally, adhesion transitions into placentation through ever-increasing development of interdigitating microvilli between trophectoderm and uterine LE that extends into the peripheral zone by day 26 of gestation.
8.4.2 Trophoblast Attachment and Adhesion
The term “implantation” is somewhat of a misnomer for the pig but is, nevertheless, used to describe the initial stages of placentation in this species. Despite differences in duration of the pre-implantation period, protracted in the pig, and type of implantation, the initial stages of implantation/placentation are common across species and are characterized as the “adhesion cascade for implantation” (Dantzer 1985; Guillomot 1995; Burghardt et al. 2002). The phases of this adhesion cascade in pigs include (1) shedding of the zona pellucida and elongation of the conceptus trophectoderm, (2) precontact and conceptus trophectoderm orientation to the uterine LE, (3) apposition of trophectoderm to uterine LE, (4) adhesion of the apical surface of trophectoderm to the apical surface of uterine LE, and (5) development of interdigitating microvilli between trophectoderm and uterine LE (Fig. 8.3). As this cascade concludes, adhesion seamlessly transitions to the progressive formation of epitheliochorial placentation that supports fetal–placental development throughout pregnancy (Johnson et al. 2014).
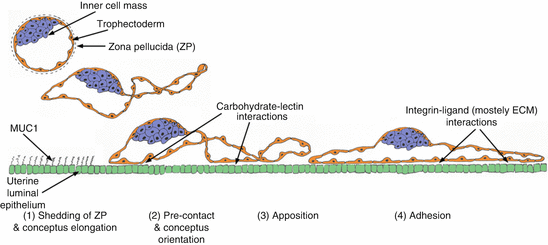
Fig. 8.3
The initial stages of implantation are common across species and are characterized as the “adhesion cascade for implantation.” The phases of this adhesion cascade in pigs include (1) elongation of conceptus trophectoderm after shedding of the zona pellucida; (2) downregulation of MUC1 at the apical surface of uterine LE to expose potential, but not yet identified, low-affinity carbohydrate–lectin binding molecules that mediate precontact and conceptus trophectoderm orientation to the uterine LE; (3) replacement of low-affinity contacts by a more stable and extensive repertoire of adhesive interactions between integrins and maternal ECM to mediate apposition and attachment of trophectoderm to uterine LE; (4) integrin receptors expressed at the apical surface of uterine LE cells bind to Arg-Gly-Asp (RGD) and non RGD amino acid sequence-containing ECM molecules and bridge to another complement of potential integrin receptors expressed at the apical surface of conceptus trophectoderm cells to mediate conceptus trophectoderm adhesion; and (5) development of interdigitating microvilli between uterine LE and trophectoderm to stabilize the trophectoderm–uterine LE interface for exchange of nutrients and gases (not illustrated in the figure)
During the peri-implantation period of pregnancy, uterine LE and conceptus trophectoderm develop adhesion competency in synchrony to initiate the adhesion cascade within a restricted period of the uterine cycle termed the “window of receptivity” (Fazleabas et al. 2004; Spencer et al. 2007; Bazer et al. 2011). Similar to other species, this window is orchestrated through the actions of progesterone and estrogen to regulate locally produced cytokines, growth factors, cell surface glycoproteins, cell surface adhesion molecules, and extracellular matrix (ECM) proteins in the pig (Johnson et al. 2009). Conceptus synthesis and release of estrogens modulate uterine gene expression (Johnson et al. 2009). The importance of estrogen to implantation of pig conceptuses is underscored by the fact that premature exposure of the pregnant uterus to estrogen on days 9 and 10 results in degeneration of all pig conceptuses by day 15 (Ashworth et al. 2006). Progesterone has a clear role in initiating the adhesion cascade for implantation in pigs; however, the mechanism by which progesterone regulates uterine epithelial cell functions remains a paradox. As long ago as 1973, studies of endocrine regulation of expression of uteroferrin secreted by the uterine epithelia of pigs revealed a requirement for long-term treatment with progesterone indicating that effects of progesterone are not mediated by a “classical” steroid receptor-type mechanism of action (Knight et al. 1973). Based on current evidence, long-term treatment with progesterone is required in order to downregulate PGR in uterine epithelia as a prerequisite to epithelial cell proliferation, gene expression, and differentiated functions. Similar to all species studied, progesterone downregulates expression of PGR in the uterine epithelia of pigs after day 10 of pregnancy, immediately prior to the time when the endometrium becomes receptive to implantation (Geisert et al. 1994; Bazer et al. 2008; Bailey et al. 2010). As previously indicated, uterine stromal cells express PGR throughout gestation, suggesting that effects of progesterone on PGR-negative uterine epithelia may be mediated through an indirect pathway whereby PGR-positive stromal cells stimulate expression of a progestamedin(s) that, in turn, modulates function of epithelial cells (Bazer et al. 2012). Although proposed progestamedins such as FGF10 and hepatocyte growth factor are expressed by the uterine stromal cells of sheep, no presumptive progestamedin has been detected in the stroma of pigs, and FGF7 is actually expressed by the uterine LE during the peri-implantation period in response to estrogen after progesterone downregulates PGR (Chen et al. 2000a, b; Ka et al. 2000). An alternative possibility is that downregulation of PGR in uterine epithelia removes an unidentified intrinsic “block” to differentiated functions in these epithelia (Spencer et al. 2004a). Regardless of the specific mechanism involved, downregulation of PGR in uterine LE is temporally associated with alterations in the expression of anti-adhesive components, mainly MUC1, an intrinsic transmembrane mucin within the glycocalyx of LE that sterically inhibits attachment of the conceptus (Brayman et al. 2004). In vivo administration of progesterone to cyclic gilts results in the loss of MUC1 from the apical surface of uterine LE (Bowen et al. 1996). It is accepted that in all mammals, initial conceptus attachment requires this loss of MUC1.
Downregulation of MUC1 exposes potential low-affinity carbohydrate ligand binding molecules including selectins and galectins and perhaps heparan sulfate proteoglycan, heparin binding EGF-like growth factors, cadherins, integrins, and CD44, which are proposed to contribute to initial attachment of conceptus trophectoderm to uterine LE (Kimber et al. 1995; Kimber and Spanswick 2000; Spencer et al. 2004b; Johnson et al. 2014). The involvement of carbohydrate–lectin interactions during the adhesion cascade of pigs has not been investigated. However, it is likely that filamentous porcine conceptuses undergo a series of attach-and-release events between carbohydrates and lectin receptors at the apical surfaces of trophectoderm and uterine LE that result in maximal apposition of these tissues, similar to the “rolling and tethering” that occur during leukocyte adhesion to the endothelium for extravasation of leukocytes (Kling et al. 1992), and proposed for the initial attachment of human blastocysts to the uterine wall (Red-Horse et al. 2004). In support of this idea, other domestic farm species, goats and sheep, exhibit prominent expression of H-type 1 antigens and glycosylation-dependent glycam 1, respectively, at the uterine–conceptus interface during implantation (Powell et al. 2000; Gray et al. 2004). These low-affinity contacts are then replaced by a more stable and extensive repertoire of adhesive interactions between integrins and maternal ECM which appear to be the major contributors to stable adhesion at implantation (Hynes 1987; Ruoslahti and Pierschbacher 1987; Aplin et al. 1994; Burghardt et al. 1997, 2002; Johnson et al. 2001; Lessey 2002). Integrins are dominant glycoproteins in many cell adhesion cascades (Kling et al. 1992). They are transmembrane glycoprotein receptors composed of non-covalently bound α and β subunits that promote cell–cell and cell–ECM adhesion, cause cytoskeletal reorganization to stabilize adhesion, and transduction signals through numerous signaling intermediates (Giancotti and Ruoslahti 1999; Albelda and Buck 1990). There are 18 α- and 8 β-subunits capable of dimerizing to form 24 different heterodimer combinations that can bind to numerous extracellular ligands including a variety of ECM proteins (Albelda and Buck 1990; Humphries et al. 2006; Gallant et al. 2005). Integrin receptors expressed at the apical surface of uterine LE cells are capable of binding to Arg-Gly-Asp (RGD) and non RGD amino acid sequence-containing ECM molecules and bridge to another complement of potential integrin receptors expressed at the apical surface of conceptus trophectoderm cells.
At present, it is known that eight integrin subunits, α1, α3, α4, α5, αv, β1, β3, and β5, are expressed at the apical surface of both uterine LE and conceptus trophectoderm. Luminal epithelial expression of α4, α5, and β1 increases during the period of maternal recognition of pregnancy, and similar increases in expression of these integrins can be stimulated in the uteri of cyclic pigs through treatment with progesterone. Further, α4, α5, αv, β1, β3, and β5 have been localized to porcine implantation sites on day 12–15 of gestation in pigs (Bowen et al. 1996). These subunits potentially give rise to the integrin receptors αvβ1, αvβ3, αvβ5, α4β1, and α5β1 that likely function as part of an adhesion cascade that serves to generate both stable adhesion between trophectoderm and LE and activation of outside-in signal transduction (Burghardt et al. 1997). Integrin-mediated adhesion results in dynamic macromolecular complexes, termed focal adhesions (FAs) which are composed of heterodimeric transmembrane integrin receptors that connect ECM proteins to the actin cytoskeleton along with a diverse array of cell signaling intermediates (Sastry and Burridge 2000; Wozniak et al. 2004; Larsen et al.. 2006). It is noteworthy that immunofluorescence staining has revealed that αv and β3 integrin subunits co-localize with altered distribution of talin (an intracellular signaling intermediate within FAs) within large aggregates at the junction between trophectoderm and uterine LE on day 20 of gestation which is approximately midway through the process of attachment for implantation and placentation in pigs (Erikson et al. 2009). The size and nature of these aggregates are reminiscent of the well-characterized FAs that form at the base of cultured cells as they attach to ECM on a rigid substrate at their basal surfaces (Sastry and Burridge 2000; Burghardt et al. 2009). Five ligands capable of engaging integrin receptors to induce assembly of focal adhesions have been characterized at sites of implantation in pigs. The inter-α-trypsin inhibitor heavy chain-like (IαIH4) protein contains a von Willebrand type A domain, a recognition site for the αvβ3 integrin receptor (Geisert et al. 1998). The latency-associated peptide (LAP) of transforming growth factor beta (TGFB) binds to αvβ1, αvβ3, and αvβ5 (Massuto et al. 2009a). Fibronectin is capable of binding α4β1, α5β1, and αvβ3 (Bowen et al. 1996). Vitronectin is a major ligand for αvβ3 (Bowen et al. 1996). Finally, osteopontin [OPN, also known as secreted phosphoprotein 1 (SPP1)] is the most promiscuous of the ligands and it interacts with αvβ1, αvβ3, αvβ5, and α4β1 (Johnson et al. 2003).
The IαIH4 is part of the kallikrein–kininogen–kinin protease system. Both IαIH4 protein expression and kallikrein enzymatic activity increase within the uterine environment during the peri-implantation period of pigs (Geisert et al. 1998; Vonnahme et al. 1999). The IαIH4 can directly engage the αvβ3 integrin receptor, as well as interact with hyaluronic acid within the ECM to aid in the initial stages of implantation. However, it is hypothesized that the primary role of IαIH4 during implantation is to act in concert with bikunin to stabilize the surface glycocalyx of the uterine LE during conceptus attachment (Hettinger et al. 2001). The LAP associates with TGFB to form an inactive homodimer called the small latent complex which remains in the cell until it is bound by another protein called latent TGFB-binding protein to form the large latent complex (LLC) that is then secreted into the ECM (Lawrence 1996). After being secreted, LAP is cleaved from the LLC by proteases to release active TGFB (Jenkins et al. 2006). In pigs, TGFB1, TGFB2, and TGFB3, as well as their receptors, TGFBRI and TGFBRII, are expressed by conceptus trophectoderm and by uterine LE between days 10 and 14 of gestation; and acting through LAP, TGFB increases fibronectin synthesis, cell adhesion to fibronectin, and the formation of FAs in a porcine trophectoderm cell line (Jaeger et al. 2005). In addition, when LAP is infused into the uteri of pregnant pigs, conceptuses failed to implant, suggesting that infused LAP competed with the endogenous LLC for binding to integrins expressed on trophectoderm (Massuto et al. 2009b). In support of this idea, aggregates of LAP, β1, β3, and β5, have been detected at the porcine conceptus trophectoderm–uterine LE interface, suggesting functional adhesion complexes that support conceptus attachment during porcine implantation (Massuto et al. 2009a). Both fibronectin and vitronectin have been detected at sites of conceptus attachment in the pig (Bowen et al. 1996), and the glycosylation variant of fibronectin, oncofetal fibronectin (oFN), is constitutively expressed by porcine conceptus trophectoderm as well as uterine LE throughout gestation (Tuo and Bazer 1996). Fibronectin and vitronectin are prototype cell adhesion proteins, and fibronectin recognizes as many as 10 different integrin receptors to generate different signal transduction functions depending upon the specific integrin receptor involved (Johansson et al. 1997; Humphries et al. 2006). It is noteworthy that the human conceptus trophoblast produces oFN so precisely at sites of trophoblast contact with endometrium that it has been referred to as “trophoblast glue” (Feinberg et al. 1994), and fibronectin is the leading candidate for conceptus adhesion molecule in rodents (Armant 2005).
The most extensively studied ECM adhesion protein for implantation in pigs is osteopontin (Johnson et al. 2003, 2014). Osteopontin is a multifunctional secreted acidic member of the small integrin-binding ligand N-linked glycoprotein (SIBLING) family of ECM proteins (Denhardt and Guo 1993; Butler et al. 1996; Sodek et al. 2000). Regarding implantation, studies have focused on the ability of OPN to support integrin-mediated cell adhesion and cell migration (Senger et al. 1994). It is likely that the expression of no other ECM protein is conserved across different species to the degree that has been observed for OPN, which is expressed abundantly within the conceptus–uterine environment of humans, mice, rabbits, sheep, goats, and pigs (Johnson et al. 1999a, b; Garlow et al. 2002; Apparao et al. 2003; Mirkin et al. 2005; Joyce et al. 2005; White et al. 2006). In pigs, expression of OPN is initially induced by conceptus estrogens in discrete regions of the uterine LE juxtaposed to the conceptus trophectoderm just prior to implantation on day 13, expands to the entire uterine LE by day 20 when firm adhesion of conceptus trophectoderm to uterine LE occurs, and remains high at this interface throughout pregnancy (Garlow et al. 2002; White et al. 2005). In vitro affinity chromatography and immunoprecipitation experiments illustrated that OPN directly binds the αvβ6 integrin heterodimer on porcine trophectoderm cells and the αvβ3 integrin heterodimer on uterine LE cells (Erikson et al. 2009). In addition, OPN binding promotes dose- and integrin-dependent attachment of trophectoderm and uterine LE cells and stimulates haptotactic trophectoderm cell migration, meaning that cells migrated directionally along a physical gradient of nonsoluble OPN (Erikson et al. 2009). Interestingly, immunofluorescence staining of tissue sections of porcine implantation sites revealed FAs containing these same integrins distributed in a similar cell-type-specific pattern to that suggested by in vitro binding to OPN. The αv integrin subunit staining revealed large aggregates at the junction between trophectoderm and uterine LE, suggesting the formation of FAs at the apical surfaces of both conceptus trophectoderm and uterine LE that facilitate conceptus attachment to the uterus for implantation. The β3 subunit, however, appeared in aggregates on the apical surface of uterine LE, but not trophectoderm, which supports results from affinity chromatography studies indicating direct in vitro binding of αvβ3 on uterine LE to OPN (Erikson et al. 2009). Finally, OPN-coated microspheres co-localize with the αv integrin subunit and talin at FAs present on the apical domain of porcine trophectoderm cells in vitro (Erikson et al. 2009). Collectively, results indicate that OPN binds integrins to stimulate integrin-mediated FA assembly, attachment, and cytoskeletal force-driven migration of conceptus trophectoderm cells to promote implantation of conceptuses in pigs.
Related posts:
 Reflections on Rodent Implantation
Implantation and Establishment of Pregnancy in Human and Nonhuman Primates
Transmembrane Mucin Expression and Function in Embryo Implantation and Placentation
Embryonic Diapause and Maternal Recognition of Pregnancy in Diapausing Mammals
The Role of Progesterone in Maternal Recognition of Pregnancy in Domestic Ruminants
The Dog: Nonconformist, Not Only in Maternal Recognition Signaling
Reflections on Rodent Implantation
Implantation and Establishment of Pregnancy in Human and Nonhuman Primates
Transmembrane Mucin Expression and Function in Embryo Implantation and Placentation
Embryonic Diapause and Maternal Recognition of Pregnancy in Diapausing Mammals
The Role of Progesterone in Maternal Recognition of Pregnancy in Domestic Ruminants
The Dog: Nonconformist, Not Only in Maternal Recognition Signaling
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


