Chapter 15 Erythrocyte and leucocyte cytochemistry
Erythrocyte cytochemistry
Siderocytes and Sideroblasts
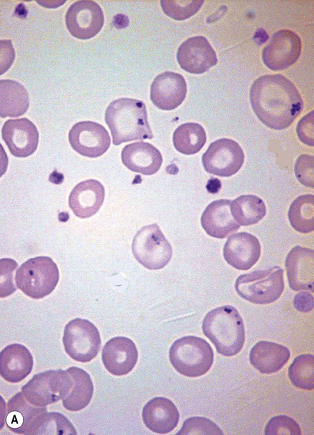
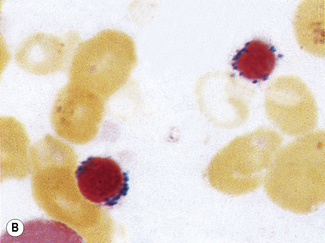
Siderocytes are red cells containing granules of non-haem iron. They were originally described by Grüneberg1 in small numbers in the blood of normal rat, mouse and human embryos and in large numbers in mice with a congenital anaemia. The granules are formed of a water-insoluble complex of ferric iron, lipid, protein and carbohydrate. This siderotic material (or haemosiderin) reacts with potassium ferrocyanide to form a blue compound, ferriferrocyanide; this reaction is the basis of a positive Prussian-blue (Perls’) reaction. The material also stains with Romanowsky dyes and then appears as basophilic granules, which have been referred to as ‘Pappenheimer bodies’ (Fig. 15.1).2 By contrast, ferritin, which is a water-soluble non-haem compound of iron with the protein apoferritin, is not detectable by Perls’ reaction. Ferritin is normally present in all cells in the body, whereas, in health, haemosiderin is mainly found in macrophages in the bone marrow, liver (Kupffer cells) and spleen. When the body is overloaded with iron, as in haemochromatosis or transfusional haemosiderosis, excess iron is also found in other tissues.
In health, siderotic granules can normally be seen, in preparations stained by Perls’ reaction, in the cytoplasm of many of the erythroblasts of human bone marrow and in marrow reticulocytes.3 However, they are not normally seen in human peripheral blood red cells. After splenectomy, siderocytes can always be found in the peripheral blood, often in large numbers. The reason for this is probably because reticulocytes, after delivery from the marrow, are normally sequestered for a time in the spleen and there they complete haem synthesis, utilizing, for this purpose, the iron stored in their cytoplasm within the siderotic granules. After splenectomy, this stage of reticulocyte maturation has to take place in the bloodstream, with the result that, even in an otherwise healthy person, a small percentage of siderocytes can then be found in the peripheral blood. The spleen is also probably able to remove large siderotic granules – as may be found in disease – from red cells by a process of pitting,4 and in its absence such granules persist in the red cells throughout their lifespan.
Method of Staining Siderotic Granules
Leave the slides in the solution for about 10 min at about 20°C. Wash well in running tap water for 20 min, rinse thoroughly in distilled water and then counterstain with 1 g/l aqueous neutral red or eosin for 10–15 s. Care must be taken to avoid contamination by iron that may have been present on the slides or in staining dishes. Prepare the glassware by soaking in 3 mol/l HCl before washing (see p. 623). For quality control, a positive bone marrow film should always be stained together with the test films.
Prussian-blue staining can be applied to films that have previously been stained by Romanowsky dyes, even after years of storage. It is advisable to let the films stand in methanol overnight to remove most of the Romanowsky stain. The film should be checked before carrying out Perls’ reaction to ensure that there is no residual blue staining that could obscure Prussian-blue staining. Sundberg and Bromann described a technique whereby films were stained first by a Romanowsky dye (Wright’s stain) and then overstained by the acid-ferrocyanide method.5 This can give beautiful pictures, but the small blue-stained iron-containing granules tend to be masked in young erythroblasts by the general basophilia of the cell cytoplasm. Hayhoe and Quaglino described a method for combined periodic acid–Schiff (PAS) and iron staining.6 This may be helpful in the investigation of abnormal erythropoiesis in which the erythroblasts give a positive PAS reaction (see p. 343). A rapid method has been described for demonstrating siderotic granules by staining with 1% bromochlorphenol blue for 1 min.7 Iron-containing granules stain dark purple.
Significance of siderocytes
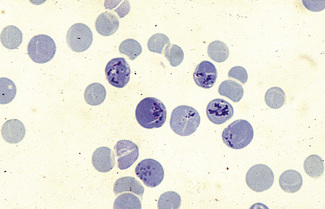
Siderocytes contain one or two (rarely many) small, unevenly distributed iron-containing granules that stain a Prussian-blue colour. There are normally a few very small scattered siderotic granules in about 40% of late erythroblasts.3 They stain faintly and may be difficult to see by light microscopy. The percentage of erythroblasts recognizable as sideroblasts is increased in haemolytic anaemias and megaloblastic anaemias and in haemochromatosis and haemosiderosis, in proportion to the degree of saturation of transferrin (i.e. to the amount of iron available). A disproportionate increase in the percentage of erythroblasts that are sideroblasts occurs when the synthesis of haemoglobin is impaired, in which case the siderotic granules are both more numerous and larger than normal (Fig. 15.2). When there is a defect in haem synthesis, the granules are deposited in mitochondria and frequently appear to be arranged in a collar around the nucleus (Fig. 15.3) giving the ‘ring sideroblasts’ characteristic of sideroblastic anaemias. In contrast, the distribution of the granules within the cell tends to be mainly normal in conditions in which globin synthesis alone is affected (e.g. in thalassaemia) or when there is iron overload.
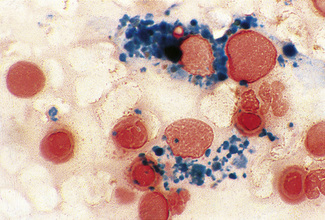
There are several types of sideroblastic anaemia. These include the congenital (hereditary) type, pyridoxine (vitamin B6) deficiency (rarely), sideroblastic anaemia caused by B6 antagonists (e.g. drugs used in antituberculosis therapy) and secondary sideroblastic anaemia in alcoholism and lead poisoning. The presence of ring sideroblasts is a defining feature of refractory anaemia with ring sideroblasts8 and refractory cytopenia with multilineage dysplasia and ring sideroblasts, two of the World Health Organization (WHO) categories of myelodysplastic syndrome (MDS). They may also occur in other categories of MDS. Ring sideroblasts are not uncommon in other haematological neoplasms, including primary myelofibrosis and acute myeloid leukaemia (AML), particularly erythroleukaemia and the WHO categories of therapy-related AML and AML with multilineage myelodysplasia. Ring sideroblasts have been defined as erythroblasts with at least five siderotic granules surrounding at least one-third of the nucleus.9,10
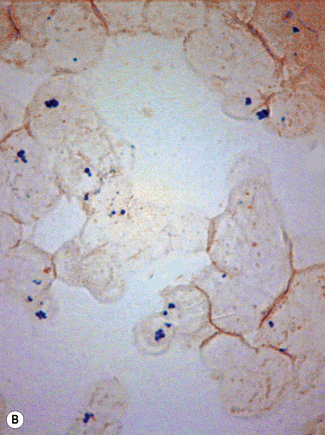
In addition to the siderotic granules within erythroblasts, haemosiderin can normally be seen in marrow films as accumulations of small granules, lying free or in macrophages in marrow fragments.11 The amount of haemosiderin will be markedly increased in patients with increased iron stores, whereas haemosiderin is absent in iron deficiency anaemia (Fig. 15.4). In practice, staining to demonstrate iron stores in marrow fragments and siderotic granules in erythroblasts is a simple and valuable diagnostic procedure and should be applied as a routine to marrow films from the initial bone marrow aspirate of each patient.
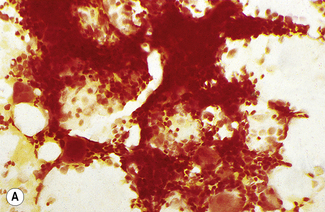
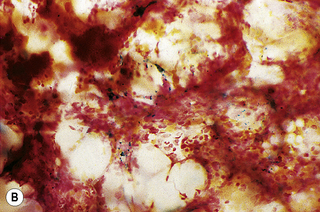
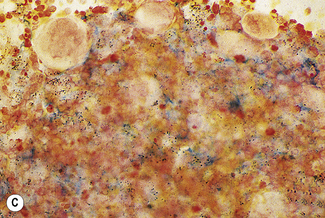

In chronic infections and in other examples of anaemia of chronic disease, the iron stores may be increased, with much siderotic material in macrophages but little or none visible in erythroblasts. Markedly excessive iron in macrophages is also a feature of thalassaemia intermedia and major and some dyserythropoietic anaemias. Conversely, absence of iron is diagnostic of iron deficiency or iron depletion (the latter term indicating the state in which storage iron is absent but anaemia is not yet evident). One study has shown that to establish the absence of stainable iron, at least seven particles must be examined, if necessary using more than one slide for this purpose.12 There is no cytochemical method of demonstrating ferritin; methods of assay are described in Chapter 9.
Haemoglobin Derivatives
Heinz Bodies in Red Cells
Heinz, in 1890, was the first to describe in detail inclusions in red cells developing as the result of the action of acetylphenylhydrazine on the blood.13 It is now known that Heinz bodies can be produced by the action on red cells of a wide range of aromatic nitro- and amino-compounds, as well as by inorganic oxidizing agents such as potassium chlorate. They also occur when one or other of the globin chains of haemoglobin is unstable. In man, the finding of Heinz bodies is a sign of either chemical poisoning, drug toxicity, glucose-6-phosphate dehydrogenase (G6PD) deficiency or the presence of an unstable haemoglobin (e.g. Hb Köln). When of chemical or drug origin, Heinz bodies are likely to be visible in red cells only if the patient has been splenectomized previously or when large doses of the chemical or drug have been taken. When they are due to an unstable haemoglobin, they are rarely visible in freshly withdrawn red cells except after splenectomy. They may nevertheless develop in vitro in the blood of patients who have not been splenectomized if the specimen is incubated for 24–48 h.14 Heinz bodies are a late sign of oxidative damage and represent an end-product of the degradation of haemoglobin. Reviews dealing with Heinz bodies include those by Jacob15 and by White.16
Demonstration of Heinz Bodies
Unstained preparations
The degradation product of an unstable haemoglobin (e.g. Hb Köln) exhibits green fluorescence when excited by blue light at 370 nm in a fluorescence microscope.17
Stained preparations
Dissolve approximately 0.5 g of methyl violet in 100 ml of 9 g/l NaCl and filter. Add 1 volume of blood (in any anticoagulant) to 4 volumes of the methyl violet solution and allow the suspension to stand for about 10 min at room temperature. Then prepare films and allow them to dry or view the suspension of cells between slide and coverglass. The Heinz bodies stain an intense purple (Fig. 15.5).
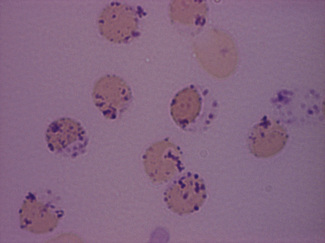
Heinz bodies also stain with other basic dyes. Brilliant green stains them well and none of the stain is taken up by the remainder of the red cell.18 Rhodanile blue (5 g/l solution in 10 g/l NaCl) stains them rapidly19 (i.e. within 2 min), at which time reticulocytes are only weakly stained. Compared with methyl violet, Heinz bodies stain less intensely with brilliant cresyl blue or New methylene blue. Nevertheless, they may be readily seen as pale blue bodies in a well-stained reticulocyte preparation, if the preparation is not counterstained.
Demonstration of Haemoglobin H Inclusions
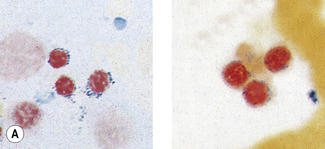
Patients with α thalassaemia, who form haemoglobin H (β4), have red cells in which multiple blue-green spherical inclusions develop on exposure to brilliant cresyl blue or New methylene blue as in reticulocyte preparations20 (Fig. 15.6). This is mainly a feature of haemoglobin H disease, but small numbers of similar cells may be seen in α thalassaemia trait, particularly, but not only, in α° thalassaemia heterozygosity.
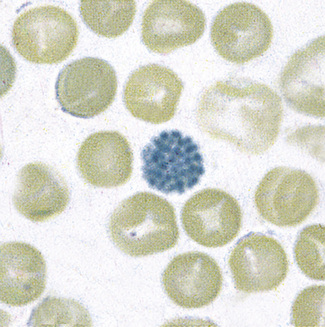
Figure 15.6 Denaturation of haemoglobin H by brilliant cresyl blue. The round bodies consist of precipitated Hb H.
Method
Mix together in a small tube, as for staining reticulocytes (see p. 33), equal volumes of fresh blood or blood collected into ethylenediaminetetra-acetic acid (EDTA) and 10 g/l brilliant cresyl blue or 20 g/l New methylene blue in iso-osmotic phosphate buffer pH 7.4. Leave the preparation at 37°C for 3 h and make films at intervals during this time. Allow the films to dry and examine them without counter-staining. Haemoglobin H precipitates as multiple pale-staining greenish-blue, almost spherical, bodies of varying size (Fig. 15.7), which can be clearly differentiated from the darker-staining reticulofilamentous material of reticulocytes (Fig. 15.8).
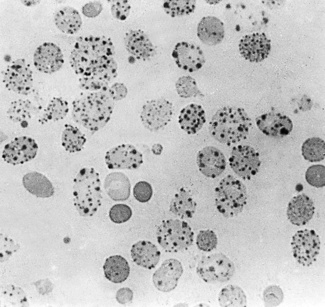
It should be noted that a haemoglobin H preparation is not recommended when precise diagnosis of the type of α thalassaemia trait is required (e.g. in antenatal diagnosis). DNA analysis is then indicated (see p. 147).
Carboxyhaemoglobin and Methaemoglobin
Carboxyhaemoglobin- and methaemoglobin-containing cells can be demonstrated cytochemically. These methods are described by Kleihauer and Betke.21 They have little practical value in modern practice.
Fetal Haemoglobin
An acid-elution cytochemical method that was introduced by Kleihauer et al.22 is a sensitive procedure to identify individual cells containing haemoglobin F even when few are present. Their detection in the maternal circulation has provided valuable information on the pathogenesis of haemolytic disease of the newborn.
The identification of cells containing haemoglobin F depends on the fact that they resist acid elution to a greater extent than do normal cells; thus, in the technique described in the following, they appear as isolated, darkly stained cells among a background of palely staining ghost cells. The occasional cells that stain to an intermediate degree are less easy to evaluate; some may be reticulocytes because these also resist acid elution to some extent. The following method, in which elution is carried out at pH 1.5, is recommended.23
Reagents
Method
Prepare fresh air-dried films. Immediately after drying, fix the films for 5 min in 80% ethanol in a Coplin jar. Then rinse the slides rapidly in water and stand them vertically on blotting paper for about 10 min to dry. Next, place the slides for 20 s in a Coplin jar containing the elution solution. Then wash the slides thoroughly in water and finally place them in the counterstain for 2 min. Rinse in tap water and allow them to dry in the air. Fetal cells stain red and adult ghost cells stain pale pink (Fig. 15.9). Films prepared (a) from a mixture of cord blood and adult blood and (b) from normal adult blood should be stained alongside the test films as positive and negative controls, respectively.
A number of modifications of the Kleihauer method have been proposed. In one, New methylene blue is incorporated in the buffer solution, the reaction time is prolonged and buffer is used for washing the films.24 The advantage of this technique is that reticulocytes stain blue, whereas cells containing haemoglobin F stain pink.
An immunofluorescent staining method has been developed based on the use of a specific antibody against haemoglobin F, which does not react with haemoglobin A.25 By using a double-labelling procedure with rhodamine-labelled antibody against γ globin and a fluorescein-labelled antibody against β globin, it is possible to detect the presence of haemoglobin F and haemoglobin A in the same cell.26
Haemoglobin S and Other Haemoglobin Variants
Immunodiffusion with specific antibodies has been used for the identification of haemoglobin S, haemoglobin A2 and haemoglobin F in red cells.27,28 An alternative method is by detection of cells after labelling the cells with fluorescein isothiocyanate (FITC).27 By a double-labelling method similar to that described earlier, it is possible to identify haemoglobin S as well as another haemoglobin in individual cells.
Leucocyte cytochemistry
Leucocyte cytochemistry encompasses the techniques used to identify diagnostically useful enzymes or other substances in the cytoplasm of haemopoietic cells. These techniques are particularly useful for the characterization of immature cells in AML and the identification of maturation abnormalities in the myelodysplastic syndromes and myeloproliferative neoplasms. There are many variations in the staining techniques, as discussed in the recommendations of an Expert Panel of the International Committee (now Council) for Standardization in Haematology.29,30 Detailed reference works discussing the theoretical and practical aspects of cytochemistry are available.31 The use of cytochemistry to characterize lymphoproliferative disorders has been largely superseded by immunological techniques (see Chapter 16). The results of cytochemical tests should always be interpreted in relation to Romanowsky stains and immunological techniques. Control blood or marrow slides should always be stained in parallel to ensure the quality of the staining. The principal uses of cytochemistry are as follows:
Myeloperoxidase
Myeloperoxidase (MPO) is located in the primary and secondary granules of neutrophils and their precursors, in eosinophil granules and in the azurophilic granules of monocytes. The MPO in eosinophil granules is cyanide resistant, whereas that in neutrophils and monocytes is cyanide sensitive. MPO splits H2O2 and in the presence of a chromogenic electron donor forms an insoluble reaction product. Various benzidine substitutes have been used, of which 3,3′-diaminobenzidine (DAB) is the preferred chromogen.33,34 The reaction product is stable, insoluble and non-diffusible. Staining can be enhanced by immersing the slides in copper sulphate or nitrate, but this is generally not required in normal diagnostic practice. Alternative non-benzidine-based techniques use 4-chloro-1-naphthol (4CN)35 or 3-amino-9-ethylcarbazole.36 The former gives very crisp staining but is soluble in some mounting media and immersion oil; the latter shows some diffusibility and does not stain as strongly as DAB.




