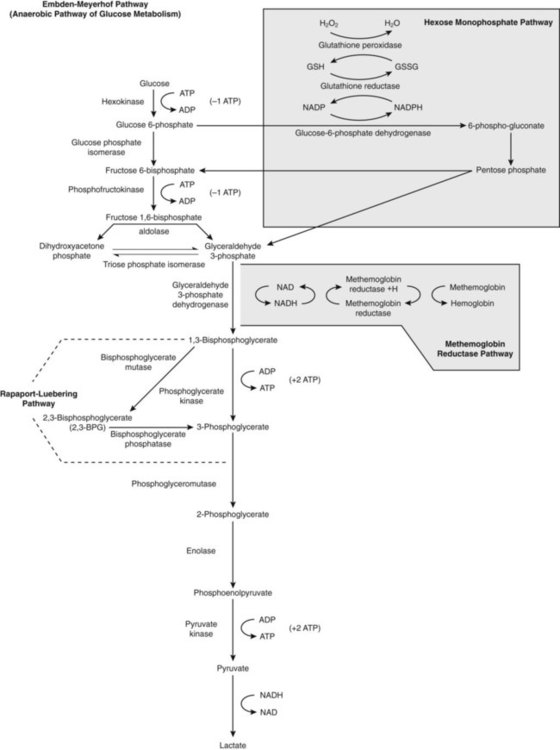
After completion of this chapter, the reader will be able to: 1. List red blood cell (RBC) processes that require energy. 2. Discuss the Embden-Meyerhof anaerobic glycolytic pathway (EMP) in the erythrocyte, with attention to adenosine triphosphate generation and consumption. 3. Name three diversion pathways from the EMP within RBCs and state the function or purpose of each. 4. Describe the role of 2,3-bisphosphoglycerate in erythrocyte metabolism and the sacrifice made to produce it. 5. Name the components related to the hexose-monophosphate pathway that act to detoxify peroxide and the type of chemical process that accomplishes the detoxification. 6. Describe the methemoglobin reductase pathway that diverts from glycolysis. 7. Explain the importance of semipermeability of biologic membranes. 8. Discuss the arrangement of lipids in the RBC membrane. 9. Discuss the function of glycolipids in the RBC membrane. 10. Explain the concept of lipid exchange between the RBC membrane and the plasma, including factors that affect the exchange. 11. Define, locate, and provide examples of transmembranous versus skeletal membrane proteins of RBCs. 12. Discuss the general structure of spectrin and its function and arrangement in the membrane. 13. Discuss how ankyrin, protein 4.1, actin, adducin, tropomodulin, dematin, and band 3 interact with α- and β-spectrin and the lipid bilayer. 14. Describe the functions of transmembranous proteins as they affect RBC function. 15. Cite the relative concentrations of sodium and potassium inside RBCs and name the structure that maintains those concentrations. 16. Name conditions that develop from abnormalities of RBC membrane constituents. Cyanosis is blue skin coloration, most obvious in whites, that occurs when the blood does not deliver enough oxygen to the tissues. It is a common sign of heart or lung disease, in which the blood fails to become oxygenated and/or is distributed improperly throughout the body. In the 1940s, Dr. James Deeny, an Irish physician, was experimenting with the use of vitamin C (ascorbic acid), a potent reducing agent, as a treatment for heart disease.1 To his disappointment, it was ineffective for nearly all patients. However, he discovered two brothers with the distinction of being truly blue men. When he treated them with vitamin C, each turned a healthy pink. Neither man was determined to have either heart or lung disease. 1. What does it mean to say that vitamin C is a reducing agent? 2. What must be happening if vitamin C was able to cure the cyanosis? 3. What is the significance of finding this condition in brothers? The red blood cell (RBC, erythrocyte) transports oxygen from the lungs to tissues and organs and helps to transport carbon dioxide to the lungs to be exhaled. The mammalian RBC has evolved to do this quite efficiently but has sacrificed greatly for this purpose by losing its nucleus.2,3 The resulting RBC shape facilitates oxygen acquisition and release. However, the cell is left without the ability to synthesize replacement proteins for vital pathways of energy production and membrane integrity, among others. As a result, its life span is limited to about 120 days because these processes inevitably fail. Mechanisms have evolved to maximize oxygen exchange during the life span of the cell and to counter the natural chemical processes that oxidize heme iron, which would leave it nonfunctional for oxygen transport. This chapter examines the energy production pathways of the RBC, discusses mechanisms for maintaining reduced heme iron and facilitating oxygen delivery, and explores the features of the RBC membrane that contribute to effective oxygen delivery for RBCs. At the time the RBC ejects its nucleus, the cellular organelles are also packaged and ejected from the cell. Without mitochondria, the RBC is left to rely on glycolysis for energy production. Fortunately, oxygen exchange and oxygen binding to heme iron do not require energy.4 Other RBC metabolic processes do require energy, however, and they are listed in Box 9-1. Loss of energy leads to RBC death. In fact, a hereditary deficiency of nearly every enzyme involved in glycolysis has been identified. A common result of these deficiencies is shortened RBC survival, known as hereditary nonspherocytic hemolytic anemia. Plasma glucose enters the RBC through a facilitated membrane transport system.5 Anaerobic glycolysis, the Embden-Meyerhof pathway (EMP; Figure 9-1), requires glucose to generate ATP, a high-energy phosphate source that is the greatest reservoir of energy in the RBC. Energy reserves like glycogen are not available in RBCs, so the RBCs rely mostly on external glucose for glycolysis-generated ATP. Via the EMP, glucose is catabolized to pyruvate, consuming two molecules of ATP per molecule of glucose and maximally generating four molecules of ATP per molecule of glucose, for a net gain of two molecules of ATP. The sequential list of biochemical intermediates involved in glucose catabolism, with corresponding enzymes, is given in Figure 9-1. Tables 9-1 through 9-3 organize this information into three phases for better comprehension. TABLE 9-1 Glucose Catabolism: First Phase TABLE 9-2 Glucose Catabolism: Second Phase TABLE 9-3 Glucose Catabolism: Third Phase The first phase of glucose catabolism involves glucose phosphorylation, isomerization, and diphosphorylation to yield fructose 1,6-bisphosphate (F-1,6-P). F-1,6-P serves as the substrate for aldolase cleavage for the final product of phase 1 glycolysis: glyceraldehyde 3-phosphate (G3P; see Figure 9-1 and Table 9-1). Hexokinase and phosphofructokinase are rate-limiting in steady-state anaerobic glycolysis, even though hexokinase has the lowest activity of all the glycolytic enzymes. The second phase of glucose catabolism converts G3P to 3-phosphoglycerate (3-PG). The substrates, enzymes, and products for this phase of glycolytic metabolism are summarized in Table 9-2. During the first reaction step, G3P is phosphorylated with a high-energy phosphate and oxidized to 1,3-bisphosphoglycerate (1,3-BPG) through the action of glyceraldehyde-3-phosphate dehydrogenase (G3PD). 1,3-BPG is dephosphorylated by phosphoglycerate kinase, which generates ATP and 3-PG. The third phase of anaerobic glucose catabolism converts 3-PG to pyruvate with the generation of ATP. Substrates, enzymes, and products are listed in Table 9-3. The product 3-PG is isomerized by phosphoglyceromutase to 2-phosphoglycerate (2-PG). Enolase then converts 2-PG to phosphoenolpyruvate (PEP). Pyruvate kinase (PK) splits off the phosphates, forming ATP and pyruvate. PK activity is allosterically modulated by increased concentrations of F-1,6-P, which enhances the affinity of PK for PEP.5 Thus when the F-1,6-P is plentiful, increased activity of PK favors pyruvate production. Pyruvic acid may diffuse from the erythrocyte or may become a substrate for lactate dehydrogenase with regeneration of the oxidixed form of nicotinamide adenine dinucleotide (NAD+). The ratio of NAD+ to the reduced form (NADH) may modify the activity of this enzyme. Aerobic or oxidative glycolysis occurs through a diversion of glucose catabolism into the HMP, also known as the pentose phosphate shunt (see Figure 9-1). The HMP detoxifies accumulated peroxide, an agent that oxidizes heme iron, proteins, and lipids, especially those containing thiol groups. Peroxide arises spontaneously from the reduction of oxygen in the cell’s aqueous environment.5 By detoxifying peroxide, the HMP extends the functional life span of the RBC. During steady-state glycolysis, 5% to 10% of G6P is diverted to the HMP. After oxidative challenge, the activity of the HMP may increase twentyfold to thirtyfold.6 The HMP catabolizes G6P to ribulose 5-phosphate and carbon dioxide by oxidizing G6P at carbon 1. The substrates, enzymes, and products of the HMP are listed in Table 9-4. TABLE 9-4 Glucose Catabolism: Hexose Monophosphate Pathway 6-PG, 6-Phosphogluconate; G6P, glucose 6-phosphate; R5P, ribulose 5-phosphate. G6PD provides the only means of generating NADPH for glutathione reduction, and in its absence erythrocytes are particularly vulnerable to oxidative damage.7 Normal G6PD activity is able to detoxify oxidative compounds and safeguard hemoglobin, sulfhydryl-containing enzymes, and membrane thiols, allowing normally functioning RBCs to carry enormous quantities of oxygen safely. However, G6PD deficiency is the most common human RBC enzyme deficiency worldwide resulting in hereditary nonspherocytic anemia (see Chapter 23). Heme iron is constantly exposed to oxygen, an oxidizing agent.8 In addition, the accumulation of peroxide oxidizes heme iron from the ferrous to the ferric state, forming methemoglobin. Although the HMP is able to prevent some hemoglobin oxidation by reducing peroxide, it is not able to reduce methemoglobin once it forms. NADPH is able to do so, but only slowly. The reduction of methemoglobin by NADPH is far more efficient in the presence of methemoglobin reductase, more properly called cytochrome b5 reductase (cytob5r). Using H+ from NADH formed when G3P is converted to 1,3-BPG, soluble cytochrome b5 reductase acts as an intermediate electron carrier, swiftly returning the oxidized iron to its ferrous, oxygen-carrying state. Cytochrome b5 reductase accounts for more than 65% of the methemoglobin-reducing capacity within the RBC.8 RBCs are biconcave and average 90 fL in volume.9 Their average surface area is 140 µm2, a 40% excess of surface area compared with a 90-fL sphere. This excess surface-to-volume ratio enables RBCs to stretch undamaged up to 2.5 times their resting diameter as they pass through narrow capillaries and through splenic pores 2 µm in diameter, a property called RBC deformability.10 The RBC plasma membrane, which is 5 µm thick, is 100 times more elastic than a comparable latex membrane, yet has tensile (lateral) strength greater than that of steel. The deformable RBC membrane provides the broad surface area and close tissue contact necessary to support the delivery of oxygen from lungs to body tissue and carbon dioxide from body tissue to lungs. RBC deformability depends not only on RBC geometry but also on relative cytoplasmic (hemoglobin) viscosity. The normal mean cell hemoglobin concentration (MCHC) ranges from 32% to 36% (see Chapter 14 and inside front cover), and as hemoglobin concentration rises, viscosity rises.11 Hemoglobin concentrations above 36% compromise deformability and shorten the RBC life span, because the more viscous cells cannot accommodate to narrow capillaries or splenic pores. As RBCs age, they lose membrane surface area while retaining hemoglobin. The hemoglobin becomes more and more concentrated, and eventually the RBC, unable to pass through the splenic pores, is destroyed by splenic macrophages. Refer to Chapter 8 for a more complete discussion of RBC senescence theories. Membrane elasticity (pliancy) is the third property that contributes to deformability. The RBC membrane consists of approximately 8% carbohydrates, 52% proteins, and 40% lipids.12 The lipid portion, equal parts of cholesterol and phospholipids, forms a bilayer universal to all animal cells (see Figure 13-9). Four main phospholipids form an impenetrable fluid barrier as their hydrophilic polar head groups are arrayed upon the membrane’s surfaces, oriented toward both the aqueous plasma and the cytoplasm, respectively.13 Their hydrophobic nonpolar acyl tails
Energy Metabolism and Membrane Physiology of the Erythrocyte
Case Study
Energy Production—Anaerobic Glycolysis
Substrates
Enzyme
Products
Glucose, ATP
Hexokinase
G6P, ADP
G6P
Glucose phosphate isomerase
F6P
F6P, ATP
Phosphofructokinase
F-1,6-P; ADP
F-1,6-P
Fructodiphosphate adolase
DHAP, G3P
Substrates
Enzyme
Product
G3P
Glyceraldehyde-3-phosphate dehydrogenase
1,3-BPG
1,3-BPG, ADP
Phosphoglycerate kinase
3-PG, ATP
1,3-BPG
Bisphosphoglyceromutase
2,3-BPG
2,3-BPG
Bisphosphoglycerate phosphatase
3-PG
Substrates
Enzyme
Product
3-PG
Monophosphoglyceromutase
2-PG
2-PG
Phosphopyruvate hydratase (enolase)
PEP
PEP, ADP
Pyruvate kinase
Pyruvate, ATP
Glycolysis Diversion Pathways (Shunts)
Hexose Monophosphate Pathway
Substrates
Enzyme
Product
G6P
Glucose-6-phosphate dehydrogenase
6-PG
6-PG
R5P

Methemoglobin Reductase Pathway
Rbc Membrane
RBC Deformability
RBC Membrane Lipids
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Energy Metabolism and Membrane Physiology of the Erythrocyte