Mammary glands are a distinguishing feature of mammals. Nursing of the young in the animal kingdom has many physiologic advantages for the mother, such as aiding postpartum uterine involution, and for the neonate, in terms of the transfer of immunity and bonding. It has become increasingly apparent that the advantages of nursing are substantial for both mother and child.
An understanding of the morphology and physiology of the breast, and the many endocrine interrelationships of both, is essential to the study of the pathophysiology of the breast and the management of benign, preneoplastic, and neoplastic disorders.
EMBRYOLOGY
During the fifth week of human fetal development, the ectodermal primitive milk streak, or “galactic band,” develops from axilla to groin on the embryonic trunk (1). The ectoderm over the thorax invaginates into the surrounding mesenchyme, with subsequent epithelial budding and branching (2). In the region of the thorax, the band develops to form a mammary ridge, whereas the remaining galactic band regresses. Incomplete regression or dispersion of the primitive galactic band leads to accessory mammary tissues, found in 2% to 6% of women in the form of accessory nipples or axillary breast tissue.
At 7 to 8 weeks’ gestation, a thickening occurs in the mammary anlage (milk hill stage), followed by invagination into the chest wall mesenchyme (disc stage) and tridimensional growth (globular stage). Further invasion of the chest wall mesenchyme results in a flattening of the ridge (cone stage) at 10 to 14 weeks’ gestation. Between 12 and 16 weeks’ gestation, mesenchymal cells differentiate into the smooth muscle of the nipple and areola. Epithelial buds develop (budding stage) and then branch to form 15 to 25 strips of epithelium (branching stage) at 16 weeks’ gestation; these strips represent the future secretory alveoli (3). The secondary mammary anlage then develops, with differentiation of the hair follicle, sebaceous gland, and sweat gland elements, but only the sweat glands develop fully at this time. Phylogenetically, the breast parenchyma is believed to develop from sweat gland tissue. In addition, apocrine glands develop to form the Montgomery glands around the nipple. The developments described thus far are independent of hormonal influences.
During the third trimester of pregnancy, placental sex hormones enter the fetal circulation and induce canalization of the branched epithelial tissues (canalization stage) (4). This process continues from the 20th to the 32nd week of gestation. At term, 15 to 25 mammary ducts have been formed, with coalescence of approximately 10 major ducts and sebaceous glands near the epidermis (5). Parenchymal differentiation occurs at 32 to 40 weeks with the development of lobuloalveolar structures that contain colostrum (end-vesicle stage). A fourfold increase in mammary gland mass occurs at this time, and the nipple-areolar complex develops and becomes pigmented. Externally the nipple is small and flattened, although rudimentary sebaceous glands and Montgomery tubercles are present. The circular smooth muscle fibers that lead to the erectile function of the nipple are developed by this stage.
In the neonate, the stimulated mammary tissue secretes colostral milk (sometimes called witch’s milk), which can be expressed from the nipple for 4 to 7 days postpartum in most neonates of either sex. At birth, the withdrawal of maternal steroids results in the secretion of neonatal prolactin. It is this hormone that stimulates newborn breast secretion. In the newborn, colostral secretion declines over a 3- to 4-week period owing to involution of the breast after withdrawal of placental hormones. During early childhood, the end vesicles become further canalized and develop into ductal structures by additional growth and branching.
After birth, the male breast undergoes minimal additional development and remains rudimentary. In the female, the breasts undergo extensive further development, which is regulated by hormones that influence reproduction. The breast has reached its major development by 20 years of age and will usually begin to undergo atrophic changes in the fifth decade of life.
DEVELOPMENTAL ABNORMALITIES
The developmental abnormalities may be unilateral or bilateral and involve both the nipple and the breast or both. These abnormalities are usually isolated to the breast, but there are reports of being associated with a variety of other abnormalities. The most common association is with upper limb and urinary tract abnormalities.
Congenital Abnormalities
Polythelia and Polymastia
The most frequently observed abnormality seen in both sexes is an accessory nipple (polythelia). Ectopic nipple tissue may be mistaken for a pigmented nevus, and it may occur at any point along the milk streak from the axilla to the groin. The reported incidence of polythelia varies greatly in the literature. In a prospective study, Mimoumi et al. (6) found the incidence of polythelia to be 2.5%. Urbani and Betti (7) evaluated the association between polythelia and kidney and urinary tract malformations. These data indicate a significantly higher frequency of kidney and urinary tract malformations in patients with polythelia. This is a controversial issue, and many studies in the literature do not find any connection between polythelia and renal anomalies (8, 9).
Rarely, accessory true mammary glands develop; these are most often located in the axilla (polymastia). During pregnancy and lactation, an accessory breast may enlarge; occasionally, if it has an associated nipple, the accessory breast may function.
Hypoplasia and Amastia
Hypoplasia is the underdevelopment of the breast; congenital absence of a breast is termed amastia. When breast tissue is lacking but a nipple is present, the condition is termed amazia. A wide range of breast abnormalities have been described and can be classified as follows (10, 11):
Unilateral hypoplasia, contralateral normal Bilateral hypoplasia with asymmetry Unilateral hyperplasia, contralateral normal Bilateral hyperplasia with asymmetry Unilateral hypoplasia, contralateral hyperplasia Unilateral hypoplasia of breast, thorax, and pectoral muscles (Poland’s syndrome)
Most of these abnormalities are not severe. The most severe deformity, amastia or marked breast hypoplasia, is associated with hypoplasia of the pectoral muscle in 90% of cases (12), but the reverse does not apply. Of women with pectoral muscle abnormalities, 92% have a normal breast (13). Congenital abnormalities of the pectoral muscle are usually manifested by the lack of the lower third of the muscle and an associated deformity of the ipsilateral rib cage. The association among absence of the pectoral muscle, chest wall deformity, and breast abnormalities was first recognized by Poland in 1841. The original description, however, did not note the concomitant abnormalities of the hand (symbrachydactyly, with hypoplasia of the middle phalanges and central skin webbing) (14), and considerable controversy has evolved concerning the validity of the eponym for this congenital syndrome (15, 16).
Athelia
The congenital absence of the nipple areolar complex is a rare entity and is usually associated with absence of the breast. This condition is typically associated with other anomalies.
Acquired Abnormalities
The most common—and avoidable—cause of amastia is iatrogenic. Injudicious biopsy of a precociously developing breast results in excision of most of the breast bud and subsequent marked deformity during puberty. The use of radiation therapy in prepubertal girls to treat either hemangioma of the breast or intrathoracic disease can also result in amastia. Traumatic injury of the developing breast, such as that caused by a severe cutaneous burn, with subsequent contracture, can also result in deformity.
NORMAL BREAST DEVELOPMENT DURING PUBERTY
Puberty in girls begins at the age of 10 to 12 years as a result of the influence of hypothalamic gonadotropin-releasing hormones secreted into the hypothalamic-pituitary portal venous system. The basophilic cells of the anterior pituitary release follicle-stimulating hormone and luteinizing hormone. Follicle-stimulating hormone causes the primordial ovarian follicles to mature into Graafian follicles, which secrete estrogens, primarily in the form of 17-estradiol. These hormones induce the growth and maturation of the breasts and genital organs (17). During the first 1 to 2 years after menarche, hypothalamic-adenohypophyseal function is unbalanced because the maturation of the primordial ovarian follicles does not result in ovulation or a luteal phase. Therefore, ovarian estrogen synthesis predominates over luteal progesterone synthesis. The physiologic effect of estrogens on the maturing breast is to stimulate longitudinal growth of ductal epithelium. Terminal ductules also form buds that precede formation of breast lobules. Simultaneously, periductal connective tissues increase in volume and elasticity, with enhanced vascularity and fat deposition. These initial changes are induced by estrogens synthesized in immature ovarian follicles, which are anovulatory; subsequently, mature follicles ovulate, and the corpus luteum releases progesterone. The relative role of these hormones is not clear. In experimental studies, estrogens alone induce a pronounced ductular increase, whereas progesterone alone does not. The two hormones together produce full ductular-lobular-alveolar development of mammary tissues (17). The marked individual variation in development of the breast makes it impossible to categorize histologic changes on the basis of age (4). Breast development by age has been described by external morphologic changes. The evolution of the breast from childhood to maturity has been divided into five phases by Tanner (18), as shown in Table 1-1.
MORPHOLOGY
Adult Breast
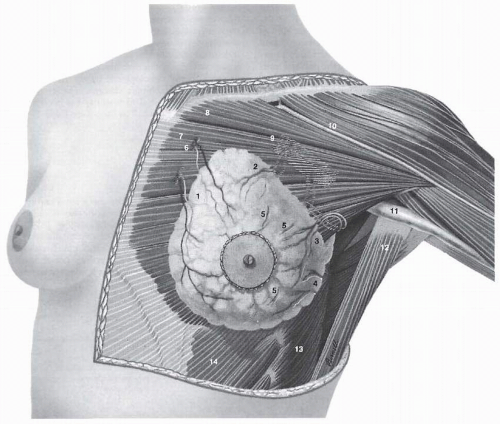
The adult breast lies between the second and sixth ribs in the vertical axis and between the sternal edge and the midaxillary line in the horizontal axis (Fig. 1-1). The average breast measures 10 to 12 cm in diameter, and its average thickness centrally is 5 to 7 cm. Breast tissue also projects into the axilla as the axillary tail of Spence. The contour of the breast varies but is usually dome-like, with a conical configuration in the nulliparous woman and a pendulous contour in the parous woman. The breast is comprised of three major structures: skin, subcutaneous tissue, and breast tissue, with the last comprising both parenchyma and stroma. The parenchyma is divided into 15 to 20 segments that converge at the nipple in a radial arrangement. The collecting ducts that drain each segment are 2 mm in diameter, with subareolar lactiferous sinuses of 5 to 8 mm in diameter. Approximately 10 major collecting milk ducts open at the nipple (5).
TABLE 1-1 Phases of Breast Development
Phase I
Age: puberty
Preadolescent elevation of the nipple with no palpable glandular tissue or areolar pigmentation.
Phase II
Age: 11.1 ± 1.1 yr
Presence of glandular tissue in the subareolar region. The nipple and breast project as a single mound from the chest wall.
Phase III
Age: 12.2 ± 1.09 yr
Increase in the amount of readily palpable glandular tissue with enlargement of the breast and increased diameter and pigmentation of the areola. The contour of the breast and nipple remains in a single plane.
Phase IV
Age: 13.1 ± 1.15 yr
Enlargement of the areola and increased areolar pigmentation. The nipple and areola form a secondary mound above the level of the breast.
Phase V
Age: 15.3 ± 1.7 yr
Final adolescent development of a smooth contour with no projection of the areola and nipple.
From Tanner JM. Wachstun und Reifung des Menschen. Stuttgart: Georg Thieme Verlag, 1962, with permission.
The nomenclature of the duct system is varied. The branching system can be named in a logical fashion, starting with the collecting ducts in the nipple and extending to the ducts that drain each alveolus, as shown in Table 1-2. Each duct drains a lobe made up of 20 to 40 lobules. Each lobule consists of 10 to 100 alveoli or tubulosaccular secretory units (5, 19). The stroma and subcutaneous tissues of the breast contain fat, connective tissue, blood vessels, nerves, and lymphatics.
The skin of the breast is thin and contains hair follicles, sebaceous glands, and eccrine sweat glands. The nipple, which is located over the fourth intercostal space in the nonpendulous breast, contains abundant sensory nerve endings, including Ruffini-like bodies and end bulbs of Krause. Moreover, sebaceous and apocrine sweat glands are present, but not hair follicles. The areola is circular and pigmented, measuring 15 to 60 mm in diameter. The Morgagni tubercles, located near the periphery of the areola, are elevations formed by openings of the ducts of the Montgomery glands. The Montgomery glands are large sebaceous glands capable of secreting milk; they represent an intermediate stage between sweat and mammary glands. Fascial tissues envelop the breast; the superficial pectoral fascia envelops the breast and is continuous with the superficial abdominal fascia of Camper. The undersurface of the breast lies on the deep pectoral fascia, covering the pectoralis major and anterior serratus muscles. Connecting these two fascial layers are fibrous bands (Cooper suspensory ligaments) that represent the “natural” means of support of the breast.
Vascular Anatomy of the Breast
The principal blood supply to the breast is derived from the internal mammary and lateral thoracic arteries. Approximately 60% of the breast, mainly the medial and central parts, is supplied by the anterior perforating branches of the internal mammary artery. Approximately 30% of the breast, mainly the upper, outer quadrant, is supplied by the lateral thoracic artery. The pectoral branch of the thoracoacromial artery; the lateral branches of the third, fourth, and fifth intercostal arteries; and the subscapular and thoracodorsal arteries all make minor contributions to the blood supply.
The principal veins involved in the venous drainage of the thoracic wall and the breast are the perforating branches of the internal thoracic vein, tributaries of the axillary vein, and perforating branches of posterior intercostal veins.
Lymphatic Drainage of the Breast
Lymph Vessels
The lymphatic drainage of the breast is of great importance in the spread of malignant disease of the breast. The subepithelial or papillary plexus of the lymphatics of the breast is confluent with the subepithelial lymphatics over the surface of the body. These valveless lymphatic vessels communicate with subdermal lymphatic vessels and merge with the Sappey subareolar plexus. The subareolar plexus receives lymphatic vessels from the nipple and areola and communicates by way of vertical lymphatic vessels equivalent to those that connect the subepithelial and subdermal plexus elsewhere (20). Lymph flows unidirectionally from the superficial to deep plexus and from the subareolar plexus through the lymphatic vessels of the lactiferous ducts to the perilobular and deep subcutaneous plexus. The periductal lymphatic vessels lie just outside the myoepithelial layer of the duct wall (21). Flow from the deep subcutaneous and intramammary lymphatic vessels moves centrifugally toward the axillary and internal mammary lymph nodes. Injection studies with radiolabeled colloid (22) have demonstrated the physiology of lymph flow and have countered the old hypothesis of centripetal flow toward the Sappey subareolar plexus (23). Approximately 3% of the lymph from the breast is estimated to flow to the internal mammary chain, whereas 97% flows to the axillary nodes (24).
New insight into lymphatic anatomy and the physiology of lymph flow has been gained from sentinel lymph node studies. It has been observed that the dermal and parenchymal lymphatics drain to the same axillary lymph nodes that are the main basin for lymph draining from the breast (25, 26, 27, 28, 29 and 30). This might be expected considering the embryology of the breast described earlier in this chapter.
FIGURE 1-1Normal anatomy of the breast and pectoralis major muscle. 1. Perforating branches from internal mammary artery and vein; 2. Pectoral branches from thoracoacromial artery and vein; 3. External mammary branch from lateral thoracic artery and vein; 4. Branches from subscapular and thoracodorsal arteries and veins; 5. Lateral branches of third, fourth, and fifth intercostal arteries and veins; 6. Internal mammary artery and veins; 7. Sternocostal head of pectoralis major muscle; 8. Clavicular head of pectoralis major muscle; 9. Axillary artery and vein; 10. Cephalic vein; 11. Axillary sheath; 12. Latissimus dorsi muscle; 13. Serratus anterior muscle; 14. External abdominal oblique muscle.
Lymphoscintigraphic studies have also shown that deeper parenchymal or retromammary lymphatics preferentially drain to the internal mammary lymph nodes when compared to intradermal or subdermal injection (31, 32, 33, 34 and 35). There has been controversy over the direction of parenchymal lymph flow in relation to the subareolar plexus. Isotope injection of technetium-99m-labeled sulfur colloid into the subareolar region results in localization of isotope in the axillary sentinel lymph node (36, 37 and 38). A detailed isotope study of subareolar injection and the lymphatic channels leading to the sentinel lymph node showed that in 90% of cases a single channel exited the areolar margin superiorly or laterally and terminated in an axillary sentinel lymph node (39). Secondary lymphatic channels exited the areola in 75% of cases. None entered the internal mammary lymph node chain.
TABLE 1-2 Nomenclature of the Breast Epithelial System
Major ducts
Terminal ducts
Collecting ducts
Extralobular
Lactiferous sinuses
Intralobular
Segmental ducts
Lobules
Subsegmental ducts
Alveoli
Terminal duct-lobular unit
Suami et al. (40) studied 24 breasts in 14 fresh human cadavers to examine the lymphatic drainage. Lymph collecting vessels were found evenly spaced at the periphery of the anterior upper torso draining radially into the axillary nodes. As identified in cross-section analysis, as these collecting vessels reached the breast some passed over and some through the breast parenchyma. Perforating lymph vessels that coursed beside the branches of the internal mammary vessels and drained into the ipsilateral internal mammary lymphatics were also found. Some of these findings are discordant with current knowledge and may explain some of the false-negative rates of sentinel lymph node biopsy.
Only gold members can continue reading. Log In or Register to continue
 Pathology of Benign Breast Disorders
Pathology of Benign Breast Disorders
 Ductal Carcinoma In Situ and Microinvasive Carcinoma
Ductal Carcinoma In Situ and Microinvasive Carcinoma
 Adjuvant Systemic Therapy: Endocrine Therapy
Adjuvant Systemic Therapy: Endocrine Therapy
 Preoperative Endocrine Therapy for Operable Breast Cancer
Preoperative Endocrine Therapy for Operable Breast Cancer



