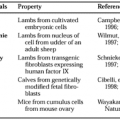
BIOLOGIC ACTIONS OF VITAMIN D
Part of “CHAPTER 54 – VITAMIN D“
INTESTINE
CALCIUM TRANSPORT
The most important bioeffect of 1,25(OH)2D3 is to increase the intestinal absorption of calcium. Although the details of the mechanisms underlying this process remain sketchy, several key steps have been characterized.33 In the absence of vitamin D, Ca2+ is able to enter the cell, but little is absorbed as a result of its sequestration in the brush border terminal web. When 1,25(OH)2D3 is given to a vitamin D–deficient animal, there is a rapid increase in Ca2+ entry over ˜30 minutes. A second phase of calcium transport peaks several hours after administration of 1,25(OH)2D3, and termination of the entire transcellular Ca2+ absorption process is not complete for several more hours. During this lag period, calbindin (see later) and other vitamin D induced proteins are synthesized and are believed to participate in the delivery of Ca2+ to the basolateral membrane, where the ion is extruded by the action of the Na+/Ca2+ exchanger. The number of functional Na+/Ca2+ exchangers is increased by vitamin-D treatment.
The best characterized vitamin D–induced protein is a cytoplasmic, 28,000-dalton calcium-binding protein (calbindin 28K) that was first identified in chick intestine.34 This protein has four high-affinity calcium-binding sites, and its dependence on vitamin D distinguishes it from calmodulin and other intracellular calcium-binding proteins. A separate 9000-dalton vitamin D–dependent calcium-binding protein (calbindin 9K), with 2 Ca2+ binding sites, is expressed in mammalian intestine. In vitamin D–deficient chickens given 1,25(OH)2D orally, both the induction of calcium transport and the appearance of calbindin in villus enterocytes are demonstrable within a few hours after dosing. As indicated earlier, calbindin is thought to play a direct role in calcium transport but it may also mediate specific calcium-dependent processes within the enterocyte. The vitamin D–dependent calbindin 28K also is expressed in kidney,35 brain,36 bone, and other tissues.34 Although the calbindins were initially viewed as vitamin D–dependent proteins, it is now clear that they are expressed in a wide array of tissues in a vitamin D–independent fashion. It has been suggested that the cal-bindins might serve as a buffer to guard against large fluctuations in intracellular calcium.
PHOSPHATE TRANSPORT
The impact of vitamin D on phosphate transport was described in the 1960s by Harrison and Harrison using an everted gut sac method. Phosphate, like calcium, is transferred across the intestinal epithelium via a multi-step process.37 An energy-dependent step is required for entry of the negatively charged phosphate ion into the cell, whereas the extrusion step occurs primarily by diffusional processes. Studies in vitamin D–deficient chicks administered32 P-phosphate suggest that vitamin D exerts effects on both the phosphate entry and exit mechanisms. The phosphate-entry step is Na+-dependent, which is reminiscent of the absorption of a number of nonelectrolytes, such as glucose and amino acids. Several isoforms of the renal Na+/phosphate co-transporters have been identified in intestinal mucosa. In addition, vitamin D induces the synthesis of a Na+/phosphate co-transporter on the basolateral membrane. A Na+/phosphate co-transporter is expressed on the basolateral membrane; however, it is unclear whether vitamin D affects the synthesis of this protein.
EFFECT ON MEMBRANE LIPID
1,25-Dihydroxyvitamin D affects the lipid composition of intestinal brush border epithelial cells. 1,25(OH)2D-induced synthesis of phosphatidylcholine occurs over a time course similar to the 1,25(OH)2D-stimulated uptake of calcium into brush border membrane vesicles.38 These observations gave rise to the concept that 1,25(OH)2D3 may exert some of its actions through nongenomic mechanisms. Consistent with this notion are studies that show rapid influxes in intracellular calcium in several cell types in vitro and acute increases in intestinal calcium uptake in ex vivo preparations of chicken intestine.39 Presumably, these activities would require 1,25(OH)2D3 binding to a membrane receptor. Such a receptor has been proposed but has not yet been identified.
SKELETON
The most familiar property of the D vitamins is their ability to cure rickets. However, normalization of blood calcium and phosphorus levels by intravenous infusion of calcium into vitamin D–deficient rats can cure rickets in the absence of circulating 1,25(OH)2D.40 Hence, the main antirachitogenic effects of 1,25(OH)2D3 rely more on its mobilization of calcium and phosphorus at the level of the intestine than on any direct effect it has on bone formation. Nonetheless, treatment of osteoblast cells with 1,25(OH)2D in vitro is associated with marked
changes in expression of osteoblast-specific genes.41 1,25(OH)2D3 decreases the rate of synthesis of collagen and citrate decar-boxylase and increases alkaline phosphatase activity and the synthesis of the noncollagenous proteins, osteocalcin, and osteopontin. The precise role of 1,25(OH)2D3 in osteoblast function remains controversial, in part because of the differences in the experimental osteoblast cell models. By contrast, the ability of 1,25(OH)2D to cause bone mineral resorption is uncontested. Studies in vitro, using neonatal calvaria or long bones pre-labeled with45 Ca, have readily demonstrated increases in cell-mediated resorption.42 1,25(OH)2D3 increases the activity of existing osteoclasts and the rate of recruitment from mononu-clear cell precursors.43 Each of these activities appears to occur indirectly, after stimulation of an osteoblast factor called osteoprotegerin, which is a soluble member of the tumor necrosis factor (TNF) receptor family (see Chap. 61).
changes in expression of osteoblast-specific genes.41 1,25(OH)2D3 decreases the rate of synthesis of collagen and citrate decar-boxylase and increases alkaline phosphatase activity and the synthesis of the noncollagenous proteins, osteocalcin, and osteopontin. The precise role of 1,25(OH)2D3 in osteoblast function remains controversial, in part because of the differences in the experimental osteoblast cell models. By contrast, the ability of 1,25(OH)2D to cause bone mineral resorption is uncontested. Studies in vitro, using neonatal calvaria or long bones pre-labeled with45 Ca, have readily demonstrated increases in cell-mediated resorption.42 1,25(OH)2D3 increases the activity of existing osteoclasts and the rate of recruitment from mononu-clear cell precursors.43 Each of these activities appears to occur indirectly, after stimulation of an osteoblast factor called osteoprotegerin, which is a soluble member of the tumor necrosis factor (TNF) receptor family (see Chap. 61).
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


