The Major Histocompatibility Complex and Its Proteins
The Major Histocompatibility Complex and Its Proteins
David H. Margulies
Kannan Natarajan
Jamie Rossjohn
James McCluskey
INTRODUCTION
The extended family of glycoproteins known as major histocompatibility complex (MHC) and MHC-like molecules is now recognized to comprise a class of receptors, usually expressed at the surface of somatic cells of vertebrates, which confer a wide range of functions in the regulation of the immune response. Their cumbersome moniker, MHC, derives from the early determination that the genes that encode these molecules control one of the fundamental manifestations of an immune response, skin graft rejection, but contemporary immunologists, virologists, and tumor biologists rarely think about these molecules with allogeneic (ie, genetically distinct) skin transplantation in mind. The name MHC is a general label, and specific immunologic functions or molecular structures invoked when a particular MHC is mentioned are dependent on biologic context. In recent years, our understanding of the structure, evolution, and molecular basis of MHC-based interactions has blossomed, and our ability to exploit engineered forms of these molecules has also expanded enormously. Our blueprint for this chapter is to give an overview of MHC-regulated immunologic function and then to summarize the explosive growth in understanding of the molecular structures of MHC molecules and their ligands. Of necessity, we discuss briefly some aspects of the cell biology and cellular maturation of MHC molecules but leave the detailed description of these essential aspects of the classic MHC molecules to the chapter on antigen presentation. Basic aspects of the genetics of MHC molecules not only descriptions of the genetic loci that encode them but also discussion of what we know of the evolution of their encoding genes will be raised.
Functional glycoproteins are molecular machines and their cellular and immunologic activities depend on the shape of these macromolecules, their surface charge and ability to interact with solvent, as well as the flexibility and relationship of their structured domains and unstructured regions. To understand molecules of the MHC in a rational functional context, we consider it crucial to describe their overall structures as well as their structural interactions with bound ligands and their binding to their cognate receptors found on T cells and natural killer (NK) cells. Despite a large and ever increasing database of MHC and MHC-like structures, a number of general questions remain unanswered. We will present some of the unsolved mysteries of the evolution and function of the MHC-like molecules and their viral homologs.
The importance of MHC molecules is underscored by two general characteristics: the enormous degree of polymorphism of genes of the Mhc and the identification of a large number of human diseases that are profoundly influenced by either defects of MHC expression or by polymorphic variants. Every student of immunology must be acquainted with the basic biology of MHC-encoded molecules, and the student’s comprehension of the regulation of the immune response, encompassing the inflammatory, NK, T-cell, and B-cell responses, will be lacking unless the function and ongoing evolution of the MHC is understood.
We stress that the evolution of the MHC is ongoing because for no other set of genetic markers is there such a large database cataloguing genetic polymorphism; as a result, it is apparent that not only are new alleles being identified, but that many of these new alleles have arisen recently. Dramatic examples of the ongoing evolution of MHC molecules are seen not only in new variants of the deoxyribonucleic acid (DNA) sequences observed by those that perform the necessary typing of Mhc genotypes for transplantation but also in surveys of the sequences of viruses, particularly the cytomegaloviruses (CMVs), which appear to have purloined host Mhc genes for immunoevasion. In sequence gazing, we observe the interplay between host and pathogen, as the pathogenic viruses, exploiting their rapid generation times, develop new variations on the MHC theme for distinct purposes.
The prototypes of MHC molecules are the MHC class I (MHC-I) and class II (MHC-II) molecules, obligate cellsurface heterodimers that bind and display self- or foreign peptides at the cell surface so that T-cell receptors (TCRs) or NK cell receptors can interact with the molecular complexes in an MHC- and peptide-dependent manner.
MHC molecules are crucial for both TCR- and NK- mediated interactions. Our efforts will be to preserve some sense of the historical development of this exciting field of study and also to focus on paradigmatic genetic, structural, and functional features that unify this extensive gene/protein family. Finally, the Mhc provides a genetic link from immune responsiveness to autoimmune disease—those well-known strong associations of particular Mhc genes to particular human diseases—and we will provide an outline of the molecular basis for such associations.
Mhc Nomenclature—Dialects of Mhc
The names of the genes and proteins that are critical to understanding the Mhc reflect the historical discovery of their functions and differ among different species. Often confusing to students of immunology, the nomenclature of the Mhc differs for different species and must deal not only with a number of distinct, but usually linked, genetic loci but also with their encoded molecules, many of which are heterodimeric glycoproteins. Because the first genes of the Mhc identified were those that encoded cell surface molecules that could be detected by antibodies or by transplantation responses, these are the ones that are referred to as Mhc genes. Now we know of more than 400 genes that map to the human or mouse Mhc, although technically they are all “Mhc” genes, the “MHC” molecules refer specifically to the MHC-I or MHC-II molecules that are related in structure and function. (Genes for several complement components and related molecules also map here and are occasionally referred to as MHC class III.) Other Mhc-encoded molecules with distinct structure and function are usually referred to by their specific names. Molecules that exhibit structural similarity with MHC-I molecules, whether they map to the Mhc or not, may be called as a group “MHC-Ib” to distinguish them from the “classical” MHC-I molecules, also referred to as “MHC-Ia.” Another subset of molecules that exhibit the MHC-I fold but that are expressed by viruses may be called “MHC-Iv” molecules.1,2
By convention, genetic loci are indicated by designations in italics, and the encoded protein products or phenotypic descriptions are shown in a standard font. The extended genetic region is referred to as the complex; thus the general term used for all species is the Mhc or MHC. The mouse Mhc is referred to as H2 (previously called H-2) because it was the second genetic locus involved in control of expression of erythrocyte antigens identified by Gorer.3,4 The Mhc in the rat is known as RT1, the human locus is known as HLA (for human leukocyte antigen), DLA for the dog, GPLA for the guinea pig, SLA for the swine, and RLA for the rabbit. For other species, based on a suggestion by Klein,5 the taxonomic name forms the basis for the designation, contributing the first two letters of the genus and the first two of the species to name the locus. Thus, we have Patr for the chimpanzee, Pan troglodytes; Gogo for the Gorilla, Gorilla gorilla; Mamu for the Rhesus macaque, Macaca mulatta; Mane for the pig-tailed macaque, Macaca nemestrina, Mafa for the cynomolgous monkey, Macaca fascicularis; and Papa for the Bonobo, Pan paniscus. A single site for curated sequences of many species of MHC molecules is the IPD-MHC Database at www.ebi.ac.uk/ipd/mhc/.6 The most important and widely studied models are those of the human, the mouse, and the rat. For the mouse and rat Mhc, the search engine on the Jackson Laboratory Web site (www.informatics.jax.org/mgihome/nomen) is the most comprehensive.
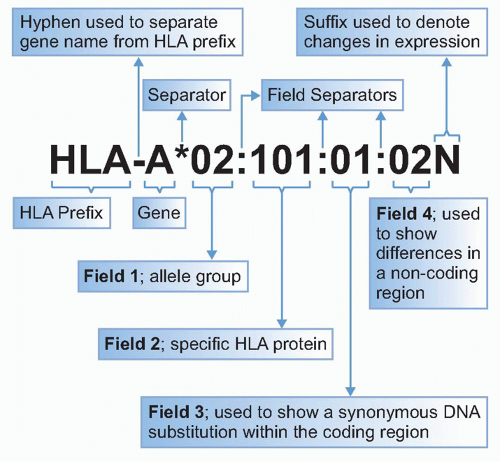
The naming of new HLA genes and alleles and their quality control are governed by the World Health Organization Nomenclature Committee for Factors of the HLA System. Software conversion tools to assist in gene/protein identification are to be found at the IMGT/HLA database (www.ebi.ac.uk/imgt/hla/dictionary.html). Regular updates on HLA nomenclature featuring new alleles can be found in the journal Tissue Antigens and via http://hla.alleles.org/nomenclature/nomenc_updates.html. The standardization of serologically defined HLA antigens has been achieved by the exchange of typing reagents and cells in the International Histocompatibility Workshops. The HLA Dictionary is a summary of HLA-A, HLA-B, HLA-C, HLA-DRB1/3/4/5, and HLA-DQB1 alleles and their association with serologically defined HLA-A, -B, -C, -DR, and -DQ antigens, respectively, available free online at the IMGT/HLA database site.7 As defined by the Nomenclature Committee (http://hla.alleles.org/nomenclature/naming.html), each HLA allele name has a unique number corresponding to up to four sets of digits separated by colons (Table 21.1 and Fig. 21.1). The digits before the first colon describe the type, which often corresponds to the serologic antigen carried by an allotype (a genetically distinguishable form of the molecule within the same species). The next set of digits lists the subtypes, numbers being assigned in the order in which DNA sequences have been determined. Longer names are assigned only when necessary. Precise designation of human genes is by a nomenclature including a number following the locus (eg, HLA-A*01:01 and HLA-DRB1*01:01). Summaries of the hundreds of human alleles that have been identified are in Tables 21.2 and 21.3. Comprehensive databases of human Mhc genes are maintained at the IMGT/HLA database (see previous link)8,9 where, at the time of this writing, more than 7000 HLA class I and class II alleles have been tabulated.
TABLE 21.1 Nomenclature of HLA Loci and Alleles
Nomenclature
Definition
HLA
The HLA region and prefix for HLA gene
HLA-DRB1
Particular HLA genetic locus
HLA-DRB1*13
Group of alleles encoding DR13 antigen or with sequence similarity to other DRB1*13 alleles
HLA-DRB1*13:01
A specific HLA allele
HLA-DRB1*13:01:02
An allele that differs by a synonymous mutation from DRB1*13:01:01 (ie, DNA sequence difference but no amino acid sequence difference)
HLA-DRB1*13:01:01:02
An allele that contains a mutation outside the coding region distinguishing it from DRB1*13:01:02
HLA-A*24:09N
A “null,” unexpressed allele
HLA-A*30:14L
An allele encoding reduced or “low” cell surface expression
HLA-A*24:02:01:02L
An allele encoding a reduced or “low” cell surface expression, where the mutation is outside the coding region
HLA-B*44:02:01:02S
An allele encoding a protein that is expressed as a “secreted” molecule only
HLA-A*32:11Q
An allele for which a previous effect on surface expression has not been confirmed and is considered “questionable”
DNA, deoxyribonucleic acid; HLA, human leukocyte antigen.
Source: Adapted from the designations summarized by the WHO Nomenclature Committee For Factors of the HLA System, Stephen G.E. Marsh, http://hla.alleles.org. Also available at the IMGT website: www.ebi.ac.uk/imgt/hla/nomenclature/index.html.
The commonly used term “haplotype” refers to the linkage of particular alleles at distinct loci that occur as a group on a parental chromosome.10 The concept of haplotype is important in typing the HLA loci in the human where the linked Mhc genes (those genes in “linkage disequilibrium”) of one chromosome of one parent will generally segregate as a group to the children. Individual haplotypes of the Mhc in the mouse are referred to by a lowercase letter superscript as H2b, H2d, or H2k. Thus, the H2k haplotype refers to the full set of linked genes, H2-Kk, H2-IAk, H2-IEk, H2-Dk, and extends to the genes of the Q and Tregions as well.11,12 (Some haplotype designations, such as H2a, refer to natural recombinants and thus have some of the linked genes from one haplotype and some from another.) A complication that demands the precise use of gene and encoded protein names is that the number of genes in a particular homologous genetic region may differ between strains or between individuals. In the mouse, although some strains have only a single gene at the D locus (H2-Db for instance), other strains may have as many as five genes in homologous region (H2-Dd, H2-D2d, H2-D3d, H2-D4d, and H2-Ld).13,14
Table 21.4 summarizes the haplotypes of common mouse strains. Included in this listing are a number of congenic inbred mouse strains, strains that contain the Mhc derived from one strain, and the remaining background genes from another. A regularly updated and comprehensive listing can be obtained at http://jaxmice.jax.org/literature/catalog/mhc_h2_haplotypes.pdf. An ongoing project, the “Collaborative Cross,” aims to establish a panel of mouse recombinant inbred lines derived from eight diverse founder mouse strains as a resource for mammalian genetics. These lines, when their breeding is completed, will provide an opportunity for mapping a wide variety of quantitative trait loci with respect to measurable phenotypes and can be expected to play an important role in studies of immune recognition.15
The Immunologic Function of MHC Molecules
MHC molecules are a molecular reflection of the health status of either the cell that synthesizes them (for MHC-I molecules) or of the local environment in which the cell resides (for MHC-II). The structure of the MHC molecule depends not only on the amino acid sequence of the two polypeptide chains (α and β for MHC-II; heavy [or α] and β2m [for the light chain] for MHC-I) that form the core of the complex but also of the variable bound peptide that forms an integral part of the trimer. The MHC molecule, governed by the sequence of the encoding structural genes for the Mhc-I heavy chain and the Mhc-II α and β chains, as well as other genes involved in antigen processing and presentation, must satisfy at least two distinct recognition functions: the binding of peptides or in some cases nonpeptidic molecules and the interaction with either T or NK cells via their respective receptors. The TCR may augment its interaction with the MHC molecule by virtue of interaction with a T cell-expressed coreceptor (cluster of differentiation [CD]8 for MHC-I and CD4 for MHC-II). Some NK receptors may also serve as coreceptors when expressed on T cells.16 The binding of peptides by an MHC-I or MHC-II molecule is the initial selective event that permits the cell expressing the MHC molecule (the antigenpresenting cell [APC] or when this cell is to be the recipient of a cytolytic signal, the target cell) to sample fragments derived either from its own proteins (for MHC-I-restricted antigen presentation) or from those proteins ingested from the immediate extracellular environment (for the case of MHC-II). The biochemical steps involved in the production of antigen fragments from large molecules are collectively known as “antigen processing,” whereas those that concern the binding of antigen fragments by MHC molecules and their display at the cell surface are known as “antigen presentation.”
Specifically, MHC-I glycoproteins gather from the cell’s biosynthetic pathway fragments of proteins derived from infecting viruses, intracellular parasites, or self-molecules, either expressed normally or in a dysregulated fashion as a result of tumorigenesis, and then display these molecular fragments, in complex with the mature MHC-I molecule, at the cell surface.17,18,19,20,21 The cell-bound MHC-I/β2m/peptide complex on the APC is exposed to the extracellular milieu and is available for interaction with either T cells or NK cells. The T cell bearing an αβ receptor recognizes the particular MHC/peptide complex by virtue of a specific physical binding interaction. Each T cell is representative of a clonal population and bears a unique TCR encoded by somatically rearranged TCR genetic elements. T cells bearing αβ receptors undergo a complex selective process in the thymus. Only a small proportion of T cells that enter the thymus ultimately reach peripheral lymphoid organs, such as lymph nodes and spleen. A particular TCR can only bind a very limited selection of MHC/peptide complexes. The recognition by T cells is considered “MHC restricted” in that only a limited set of MHC molecules can bind a particular TCR and is also termed “antigen specific” in that a particular T cell identifies a particular peptide. For any given T-cell clone, single amino acid substitutions of either the MHC or the peptide may severely diminish, obliterate, or even augment the functional interaction of the TCR with the MHC-II/peptide complex. The MHC-I system draws its spectrum of peptides from proteins in the cytosol that are degraded by the proteolytic proteasome complex to peptides that are transported from the cytosol to the endoplasmic reticulum (ER) with the aid of the intrinsic membrane peptide transporter, the transporter associated with antigen processing (TAP), are then trimmed at their amino termini22 and are cooperatively folded as an intrinsic component of the newly synthesized MHC-I molecule.23 Interactions with the peptide-loading complex (PLC), which includes TAP, the chaperone tapasin,24 and Erp57,25 are crucial to MHC-I loading. MHC-I molecules are unique among proteins in that their three-dimensional structure and thermal stability are exquisitely dependent on the heavy/light chain heterodimer being bound by an appropriate peptide.26,27 In mutant cells that lack the necessary apparatus for generating and transporting peptides to the ER where peptide loading takes place, MHC-I molecules are expressed poorly and are inherently unstable.28 MHC-II/peptide loading is controlled in part by the multifunctional chaperone/groove protector, Ii, as well as the important catalytic machinery of the endosomes, molecules known as HLA-DM and -DO in the human, and H2-M and H2-O in the mouse.24 Although there remains some controversy as to the precise site of interaction of HLA-DM with HLA-DR, evidence supports the view that this interaction catalyzes peptide interchange, leading to the selection of high-affinity antigenic peptides.29
aThis table is based on a listing of alleles maintained by the World Health Organization Nomenclature Committee for Factors of the HLA System, as of January 2012. All new and confirmatory sequences are generally submitted directly to the committee via the IMGT/HLA database using the sequence submission tool provided. The IMGT/HLA database may be accessed at www.ebi.ac.uk/imgt/hla. Serologic assignment of HLA-DR molecules is largely determined by the DRB1 gene product, whereas assignment of DQ molecules reflects serologic contributions from both DQA1 and DQB1 gene products.
HLA, human leukocyte antigen.
TABLE 21.4 Commonly Used Mouse Strains: H-2 Haplotypesa
H-2 Complex
Strain
Haplotype
KK
Ab
Aa
Eb
Ea
D
Q
T
Common strains
129/J
bc
b
b
b
b
–
b
b
f
AKR/J
k
k
k
k
k
k
k
b
b
ASW/Sn
s
s
s
s
s
–
s
b
b
BALB/c
d
d
d
d
d
d
d
b
c
C3H/HeJ
k
k
k
k
k
k
k
b
b
CBA/J
k
k
k
k
k
k
k
b
b
C57BL/6
b
b
b
b
b
–
b
b
b
C57BL/10
b
b
b
b
b
–
b
b
b
C57BR
k
k
k
k
k
k
k
a
a
DBA/2J
d
d
d
d
d
d
d
b
c
NOD/LtJ
g7
d
g7
d
–
–
b
NON/LtJ
nb1
b
nb1
?
k
k
b
NZB/BINJ
d2
d
d
d
d
d
d
a
a
NZW/LacJ
z
u
u
u
u
u
z
P/J
p
p
p
p
p
p
p
a
e
PL/J
u
u
u
u
u
u
d
RIII
r
r
r
r
r
r
r
c(r)
b
SJL
s2
s
s
s
s
–
s
a
a
Congenic strains
B10.BR
k2
k
k
k
k
k
k
B10.D2
d
d
d
d
d
d
d
B10.S
s
s
s
s
s
–
s
BALB.B
b
b
b
b
b
–
b
BALB.K
k
k
k
k
k
k
k
C3H.SW
b
b
b
b
b
–
b
Recombinant strains
A
a
k
k
k
k
k
k
A.TL
tl
s
k
k
k
k
d
B10.A
a
k
k
k
k
k
d
B10.A(1R)
h1
k
k
k
k
k
b
B10.A(2R)
h2
k
k
k
k
k
b
B10.A(3R)
I3
b
b
b
b/k
k
d
B10.A(4R)
h4
k
k
k
k/b
–
b
B10.A(5R)
I5
b
b
b
b/k
k
d
B10.T(6R)
y2
q
q
q
q
–
d
B10.S(7R)
t2
s
s
s
s
–
d
B10.S(8R)
as1
k
k
k
k/s
–
s
B10.S(9R)
t4
s
s
s
s/k
k
d
B10.HTT
t3
s
s
s
s/k
k
d
*Adapted from Kruisbeek561 and www.imgt.org/IMGTrepertoireMHC/Polymorphism/haplotypes/mouse/MHC/Mu_haplotypes.html#polymorphism. A dash indicates abnormal gene expression, although precise mechanism may differ in different strain. Blanks indicate insufficient data for characterization. Additional strains may be identified at http://jaxmice.jax.org/findmice/index.html.
NOD, nonobese diabetic.
MHC-II molecules, in contrast to MHC-I, are expressed on a more limited set of somatic cells—B cells, macrophages, dendritic cells, activated but not resting T cells in the human—and have a somewhat more specific function in peptide selection and presentation. In general, they bind peptides derived from the degradation of proteins ingested by the APC, and they sort their MHC-II molecules into cellular compartments where the degraded peptides are generated and catalytically transferred to the binding site of the MHC-II. The MHC-II antigen-presentation pathway is based on the initial assembly of the MHC-II αβ heterodimer with a dual function molecule, the invariant chain (Ii), which serves as both a chaperone to direct the αβ heterodimer to an endosomal, acidic, protein processing location where it encounters antigenic peptides and also serves to protect the antigen-binding site of the MHC-II molecule so that it preferentially will be loaded with antigenic peptides in this location.30,31,32,33 The loading of the MHC-II molecule with antigenic peptide, a process dependent on the release of the Ii derived “CLIP” peptide, in part dependent on the MHC-II-like molecule, HLA-DM in the human,34,35 H2-M in the mouse,36 then leads to the cell surface expression of MHC-II/peptide complexes. The MHC-II-recognizing T cells then secrete cytokines and may also be induced to proliferate or to undergo programmed cell death. Such MHC-restricted cytokine production that facilitates and augments the recruitment of additional inflammatory cells as well as APCs and antibody-producing cells is a contemporary explanation for what was historically referred to as “T-cell help.” Under some physiologic circumstances, particularly during viral infection, antigens incorporated into antigen-presenting dendritic cells via an “outside in” pathway may alternatively enter the MHC-I presentation pathway. This phenomenon is known as “cross-presentation.”37,38
MHC molecules perform a crucial role in the thymus by shaping the TCR repertoire as T-cell precursors mature into cells that eventually emigrate from the thymus and populate the spleen and other secondary lymphoid organs. In a complex, multistep process termed “thymic education,”39 developing T cells expressing randomly rearranged TCR αβ receptors are first subjected to positive selection on MHC-I and MHC-II molecules expressed by cells of the thymic cortex in order to select for further maturation of only those cells capable of recognizing peptides in the context of self-MHC molecules. Positive selection may be viewed as setting the minimum threshold for T-cell activation. T cells surviving positive selection then migrate to the thymic medulla where those T cells with higher-than-average affinity for self-MHC-II/peptide complexes, and which therefore pose the threat of autoimmune disease should they be allowed to emigrate to secondary lymphoid organs, are deleted by negative selection. MHC-I and MHC-II molecules thus play a critical role in shaping the peripheral TCR repertoire. Indeed, mice engineered to lack MHC-I or MHC-II expression are also deficient in T cells.36,40
Distinct from the recognition of MHC molecules by TCRs, a number of NK-cell receptors, both activating and inhibitory, bind MHC-I molecules, and several NK receptors interact with MHC-I-like molecules encoded by CMVs.2,41 In general, the NK/MHC-I interaction, as compared to the TCR/MHC interaction, shows considerably less peptide specificity, although the interaction is peptide dependent, and in some cases may exhibit clear-cut peptide preferences.42,43,44 The functional purpose of the MHC-I or MHC-I-like molecule in NK-cell recognition appears to be more subtle than that in T-cell recognition. The NK cell is tuned to a balance of inhibitory and activating signals conveyed to it via MHC interaction. In its resting state, the inhibitory signals predominate. MHC-I is a sensor of the biosynthetic and metabolic state of the cell in which it is synthesized. When the level of MHC-I is dysregulated by tumorigenesis or viral infection, this change can be detected by the NK cell. This ability of the NK cell to sense altered levels of MHC-I on target cells is the basis of the “missing self hypothesis,” which explains that NK cells detect and lyse those cells defective in MHC-I expression due to the loss of the inhibitory signal that results from engagement of NK receptors by MHC-I.45,46,47 The prototype NK receptor in the mouse is the NK inhibitory receptor, Ly49A, a C-type lectin-like molecule that signals its interaction with a normally expressed MHC-I molecule, such as H2-Dd.48 Distinct clones of NK cells differ in the combinatorial expression of different NK receptors that have different MHC preferences. Thus, in the mouse, each distinct NK clone may express a different combination of NK inhibitory receptors such as Ly49A, Ly49C, Ly49G2, and Ly49I.49 Because each inhibitory molecule may exhibit slight differences in its MHC-I and/or peptide preference and specificity, this kind of combinatorial expression of NK activity offers a breadth of specificity toward different potential target cells. Recently, it has been shown that developing NK cells within the thymus are also subject to an education process, known as “licensing,” which prevents improper activation of their cytolytic function in peripheral tissues.50,51,52 NK cells that develop in MHC-I-deficient humans and mice fail to kill target cells that lack MHC-I expression. Such targets are readily killed by NK cells that develop in MHC-I-sufficient thymi. Human NK cells seem to go through a similar selective/educating process, but the relevant NK receptors on human cells are generally of the killer cell immunoreceptor (KIR) inhibitory receptor family.53
In addition to showing preference for distinct pathways of antigen presentation, the MHC-I and MHC-II molecules also show preferential restriction to T cells of the CD8- or CD4-bearing lineages. This results from the interaction of CD8 with the nonpolymorphic α3 domain of MHC-I molecules,54,55,56,57,58 whereas CD4 binds to membrane proximal domains of MHC-II.59,60,61,62,63,64 The CD8 and CD4 molecules serve as “coreceptors” on the surface of T lymphocytes, providing both adhesion (increase in avidity) and specific activating signals, mediated through the kinase, lck, which modulate the avidity of the T cell in a time-dependent manner.65,66,67,68 Additional complexities arise from the interaction of CD8 with MHC-I. CD8 is expressed as either the CD8αα homodimer or the CD8αβ heterodimer, which are expressed in a developmentally regulated fashion and seem to have distinct functions.69 The numerous interactions of MHC molecules with other cellular components as well as with the wide variety of peptides and of various immunologic receptors reflect the robust potential of the MHC structure as a molecular sensor and as a master regulator of immune responses. These molecular interactions then read out in different cell trafficking and signaling functions.
THE MAJOR HISTOCOMPATIBILITY COMPLEX
Mhc Genetics
The Mhc is an extended region of the genome that spans some 4 million basepairs (Mb) on the short arm of human chromosome 6 in the region 6p21.3. The Mhc in the human may be considered to map from HLA-F to the gene encoding the MHC chaperone tapasin (TAP binding protein), a distance of some 2 Mb, but the “extended” human Mhc (also known as the xMHC) covers some 7.6 Mb from the HIST1H2AA locus to SYNGAP1.70,71,72 In the mouse, the Mhc encompasses almost 3.5 Mb on chromosome 17, extending from the H2-Ke1 gene at basepair position 34056423 to the H2-M2 gene at basepair position 37620474 (www.imgt.org/IMGTrepertoireMHC/LocusGenes/index.php?repertoire=listIG_TR&species=mouse&group=MHC). Although Mhc genes were among the first to be mapped here, it is now clear that a large number of genes with function unrelated to immune recognition also reside in this region. The interested reader is referred to the regularly updated maps and linkages available at various Web sites including www.ebi.ac.uk/imgt/hla, and the MHC haplotype project at www.sanger.ac.uk/HGP/Chr6/.
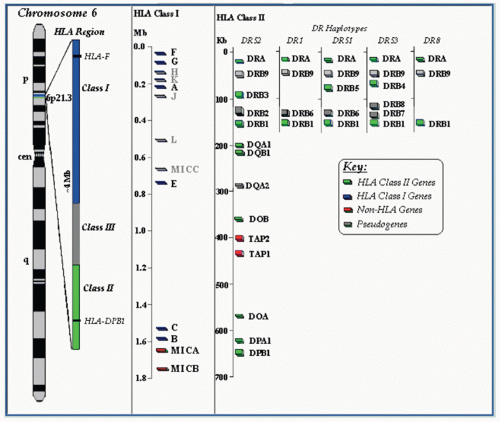
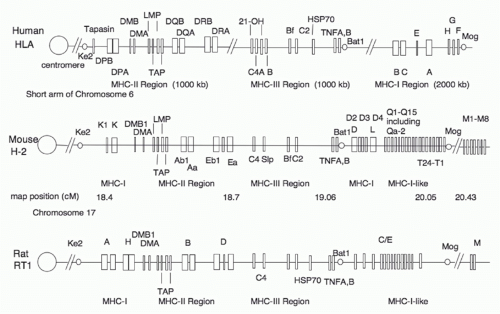
A schematic of the human Mhc is shown in Figure 21.2. For a rough guide, we also provide a simple illustrative map comparing human, mouse, and rat Mhc regions (Fig. 21.3) and refer the reader to the National Center for Biotechnology Information homology Web site: www.ncbi.nlm.nih.gov/projects/homology/maps/ and National Center for Biotechnology Information Web viewer for access to detailed maps and sequences.
FIG. 21.2. Map of the Human MHC (HLA Region) on the Short Arm of Chromosome 6. A variety of online tools for seeking and manipulating MHC sequences can be found at www.ncbi.nlm.nih.gov/gv/mhc/main.cgi?cmd=init.
The human Mhc map reveals clusters of genes grouped roughly into an Mhc-II region covering about 1000 kb, an Mhc-III of about 1000 kb, and an Mhc class I region spanning 2000 kb (see Fig. 21.2). HLA-DP genes (DPA encoding the α chain and DPB encoding the β chain) are proximal to the centromere on the short arm of the chromosome and are linked to the genes encoding the related HLA-DM molecule (DMB and DMA). Between these and the DQ genes lie LMP (for low molecular weight proteins73,74,75,76) and TAP77,78,79,80,81 genes that encode molecules that are involved in peptide generation in the cytosol and peptide transport across the ER membrane, respectively. Low molecular weight proteins are subunits of the catalytic proteolytic proteasome complex that regulate the specificity of cleavage of proteins, and thus modulate the repertoire of peptides available for MHC-I-restricted antigen presentation.82,83,84,85 The TAP genes encode a two-chain intrinsic membrane protein that resides in the ER of all cells and functions as an adenosine triphosphate-dependent transporter that pumps peptides generated in the cytosol into the lumen of the ER.86,87 The selective transport of cytoplasmically generated peptides by different TAP proteins in the rat demonstrates that the spectrum of MHC/peptide complexes expressed at the cell surface can be significantly altered by differences in the antigen presentation pathway,88,89,90 although there is little evidence for this phenomenon in humans.91 Nevertheless, TAP deficiency syndromes in humans92,93,94,95 emphasize the importance of peptide delivery in the antigen presentation pathway for normal immune function.
FIG. 21.3. Comparative Maps of Human, Mouse, and RatMhcRegions. This schematic map is not complete nor drawn to scale and is derived from maps available at www.ncbi.nlm.nih.gov/projects/homology/maps/.
The major Mhc-II genes of the human are HLA-DRA and HLA-DRB that encode the chains that form the HLA-DR molecule, a major antigen-presentation element. Genetic mapping of the human DRB region indicates that several alternative arrangements of DRB loci explain the varied serotypes and genotypes observed among different individuals (see Fig. 21.2). In particular, note that several HLA-DR “haplotypes,” originally defined serologically as DR52, DR1, DR51, DR53, and DR8, differ not only in particular allelic genes at the DR locus but also in the number and precise location of several of the DRB genes and pseudogenes. The Mhc-III region is important in immunologic terms for several reasons—the structural genes for several complement components map here, as well as the structural genes for 21-hydroxylase (CYP21A2),96,97 an enzyme critical in the biosynthesis of glucocorticoids, a deficiency of which can lead to the genetic disease congenital adrenal hyperplasia. Also located in the Mhc-III region are the structural genes for tumor necrosis factor (TNF) A (also known as lymphotoxin α) and B, which are cytokines made by activated T cells.98,99,100
The more distal region of the Mhc encodes other MHC molecules. In humans, the cluster of the major Mhc-I genes lies here, spanning two Mb, including the genes encoding HLA-B, HLA-C, HLA-E, and HLA-A, as well as HLA-H, HLA-G, and HLA-F. HLA-A, HLA-B, and HLA-C are the major MHC-I molecules of man. (A summary of these is in Table 21.2.) Serologic identification of HLA-C molecules has been difficult and imprecise. However, HLA-C molecules interact directly with NK receptors of the KIR2D family. Direct binding studies have analyzed the kinetics of the interaction of the KIR2 immunoglobulin (Ig)-like NK receptors,101,102,103 and three-dimensional structures of KIR2DL2 in complex with HLA-Cw3 and KIR2DL1 complexed with HLA-Cw4 have been published.104,105 Recently, the structure of a KIR3DL1 molecule in complex with HLA-B*57:01 emphasizes the unique nature of different KIR/MHC interactions.106 The precise functions of HLA-E, HLA-F, and HLA-G are not yet clear. HLA-E and its murine analog Qa-1 bind hydrophobic leader peptides derived from some MHC-I molecules, forming a complex recognized by the C-type lectin-like NK receptor CD94/NKG2.107,108,109,110,111 This implies an important function for HLA-E because these molecules are expressed on placental trophoblast cells and would be expected to bind the inhibitory NK receptor, CD94/NKG2A, preventing NK-mediated rejection of the fetus.112 HLA-E may serve as a recognition element for some T cells as well, so it also seems capable of a classical role.113,114 Additional evidence now supports an antigen presentation function of HLA-F and HLA-G,115,116 and the tissue restricted expression of HLA-G as well as the observation that a soluble form leads to apoptosis of CD8+ T cells suggest that this molecule may be involved in the mother’s immunologic tolerance of the fetus.117HLA-H is a pseudogene mapping to this region.118 This should not be confused with the more distantly related HLA-HFE, an Mhc-Ib gene erroneously called HLA-H by some authors,118,119 which controls hereditary hemochromatosis by virtue of the interaction of its encoded protein with the transferrin receptor.120,121,122,123,124,125 The observations that extravillous trophoblast cells express an unusual combination of HLA-C, HLA-E, and HLA-G molecules; that uterine NK cells express NK inhibitory receptor (KIR) molecules known to interact with HLA-C allelic products; and that complex genotypes predispose to preeclampsia raise the possibility that important aspects of fetal rejection and clinical infertility may be related to MHC recognition by NK cells in the placenta.126
Comparison of the mouse, rat, and human Mhc maps (Fig. 21.3) reveals several interesting differences.5,127,128,129 The Mhc genes proximal to the centromere of the mouse and rat belong to the Mhc-I family, rather than to the Mhc-II family, as they do in the human. This mapping has suggested that an intrachromosomal recombination event that occurred in some common rodent ancestor relocated some of the Mhc-I genes from a more distal location to the proximal site.130 Inspection of the current human, mouse, and rat maps clearly indicates similarities in the relative locations and organization of Mhc-II, Mhc-III, and the distal Mhc-I genes.128 Various genetic expansions and contractions131 are obvious as well. In particular, the mouse Q and T regions have expanded the pool of Mhc-I genes, which are relatively few in the human and the rat. Early studies of congenic mouse strains mapped multiple genes to the Q and T regions,132,133,134 and recent evidence suggests significant differences in the number of genes of this region in different strains. The mouse has some MHC-Ib genes that seem to be relatively unique in function. In particular, the H2-M3 gene, which maps distal to the Q and T regions, encodes a protein that exhibits a preference for binding peptides that have N-formyl amino terminal modifications. This antigen presentation function may be geared to bacterial, protozoal, and mitochondrial antigens.135,136,137 Rat homologues of the mouse H2-M3 and H2-M2 genes have also been identified.138,139
Mhc Polymorphism
The Mhc’s function in immune responsiveness is also reflected in its genetic polymorphism. Polymorphism is the presence at any given time of a larger than expected number of genetic variants in a population. As populations change and evolve, we expect that genetic variants should arise but because of the selection exerted on most gene products, relatively few of these genetic variants will persist. A genetic locus that exhibits variant alleles at a frequency of more than 1% is considered “polymorphic.”5 A genetic locus that is relatively invariant is often referred to as “monomorphic,” even if more than one allele is known. HLA genes exhibit a high degree of polymorphism; a number of different mechanisms may contribute to the generation and maintenance of polymorphism. Among these are the selective advantage of a heterozygous pool of antigen-presenting elements in a given individual that might allow the binding and presentation of antigenic peptides derived from a wide variety of environmental pathogens. Limited polymorphism would make the entire population susceptible to a chance infectious agent for which all individuals would be unable to respond, whereas widespread polymorphism would be expected to allow the APCs of at least a proportion of the population to effectively bind and present antigens derived from invading pathogens.140 Although such a view was originally based on HLA molecules as presenting elements for pathogen-derived peptide fragments to T cells and their antigen-specific TCRs, studies suggest an additional role for Mhc-I-related resistance to viral infection via NK cell-mediated recognition141,142 and altered antigen presentation pathway.143 Additionally, the presence of a high degree of polymorphism also implies that there is little or no selective disadvantage to the expression of new variant MHC molecules.
The human Mhc-I and Mhc-II genes are clearly polymorphic, with more than 5000 alleles at the Mhc-I loci and some 2000 alleles at the Mhc-II loci known (see Tables 21.2 and 21.3) (www.ebi.ac.uk/imgt/hla/index.html).8,9 In experimental animals, it is more difficult to demonstrate polymorphism in terms of population genetics, although typing of wild mice has confirmed the existence of natural polymorphism predicted from the analysis of inbred strains and their mutants.144,145 The polymorphism of Mhc-I and Mhc-II genes, so evident in man and mouse, has also been documented in analyses of cichlid fishes—animals that diverged from the line leading to mammals at least several hundred million years ago.146,147,148,149
Genetic Mechanisms for Mhc Evolution
As both an extended genetic region and a group of genes with many belonging to the Ig supergene family,150,151 the Mhc has served as a prototype for elucidating mechanisms that contribute to the evolution of a multigene family and that add to the polymorphism that is such a dominant characteristic of the classical MHC molecules.152 The analysis of mutations in the mouse, mostly those of Mhc-I genes, has led to the understanding of the mechanisms that give rise to polymorphism. Both induced and spontaneous mutations affecting skin graft acceptance or rejection have been identified, and many of these have been mapped to the Mhc. Gross recombinational events have been documented in the Mhc,153,154 as well as more subtle mutations, many of which are multiple amino acid substitutions in a relatively small part of the protein that seem to derive from nonreciprocal recombinational events. Such recombination that occurs over short sequences is known as “gene conversion” because of its similarity to the phenomenon that occurs in yeast.153,154,155,156,157,158,159,160,161 Convincing evidence for interlocus gene conversion has been documented in for the Mhc-II genes of teleost fish162 and by deep sequencing of Mhc-I genes in recently founded bird populations.163
Some of the polymorphisms of Mhc genes that have been identified clearly reflect point mutations.164 Structural studies have shown that the profound immunologic effects of mutations of the H2 genes H2Kbm1 and H2Kbm8 result from minimal detectable changes that may affect thermostability.165 In addition to such mouse mutants, a number of somatic cell variants and mutants, some due to major deletions or regulatory defects, others clearly point mutants of structural genes, have been described.166
The Mhc and Transplantation
Although the early description of the genes of the Mhc was based on identification of loci involved in tumor and allograft rejection, and although these genes clearly play a role in such complex phenomena, a contemporary understanding of the function of Mhc genes in immunology requires little understanding of the rules of transplantation. The early history of transplantation is chronicled extensively in several books5,167 and reflects a developing interest in tumor immunology and congenic mouse strains. The mouse has served as the model system for understanding the genetic underpinnings of skin, tumor, and organ transplantation, so a brief description of some relevant principles is in order. Comprehensive manuals and reviews are available.168 Propagation of a mouse strain by repeated matings of brothers and sisters leads to the establishment of an inbred strain, a group of animals that is genetically identical at all loci. More complete descriptions of the process by which brother-sister mating leads to homozygosity at all loci are given elsewhere.5,167
“Congenic” mouse strains, also known as “congenic resistant” or “CR” strains, are those derived by first crossing two inbred strains that differ in a histocompatibility phenotype such as resistance to a transplantable tumor or ability to reject a skin graft. These are then successively backcrossed to one parental strain, and the resistance phenotype is preserved. Following at least 10 backcross generations (N10), a point at which (1/2)9 = 0.002 of the genes of the selected strain should be present, the new strain is propagated by brother-sister mating. Several relevant inbred and congenic mouse strains are listed in Table 21.4 along with their H2 designations.
The early rules of transplantation were determined by observation of the ability of either transplantable tumors or allografts (usually from skin) to survive in a particular inbred mouse strain host. The graft rejection phenomenon is an extremely sensitive and specific bioassay that permits the detection of genetic differences as small as a single amino acid in an MHC protein. It has been particularly valuable in assessing spontaneous and induced mutants (see previous discussion) and remains the absolute experimental discriminator of “histocompatibility.”
In addition to the Mhc genes, we should not overlook the genes that encode minor histocompatibility antigens. In the mouse, these were originally identified as genetic loci responsible for graft rejection after extended periods of time. More recently, several minor histocompatibility loci have been identified as those that encode polymorphic proteins that give rise to peptides presented by MHC molecules,169,170,171,172,173 and we now can understand the complexities of transplantation tolerance in terms not only of Mhc genes but also in terms of numerous proteins that may give rise to variant peptides for T-cell recognition. Not only can peptides derived from polymorphic genes throughout the genome serve as minor histocompatibility antigens but also defective translation products, or peptides resulting from transcription of introns or noncoding strands of DNA may also produce immunologically significant peptides, which, bound by self-MHC molecules may stimulate T cells.174,175 Whether such defective, newly synthesized proteins serve as a major source of MHC-I-bound peptides remains controversial.176 Minor antigens that are confined to hematopoietic cells can be recognized as targets by antitumor cytolytic cells and may explain the lower incidence of relapse in hematopoietic stem cell (HSC) transplant recipients who experience graft-versus-host disease (GVHD).177
The Mhc and Clinical Transplantation
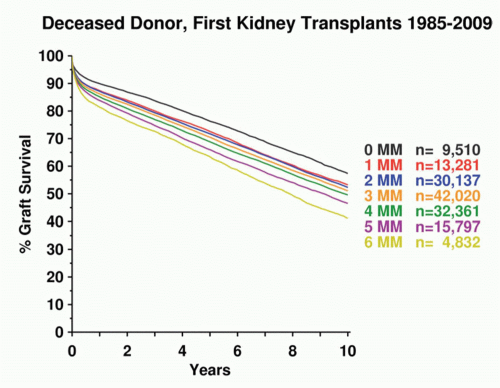
Processed foreign antigen complexed to HLA class I or class II molecules is recognized by a specific clonally distributed TCR for antigen on the surface of T lymphocytes. The T cell bearing an αβ receptor is capable of recognizing the unitary structure of the HLA molecule itself coordinately with the exposed parts of the peptide antigen. Corecognition of HLA and peptide antigen means that TCRs are highly specific and genetically restricted to recognizing HLA molecules of the individual from which they were derived. Thus, a killer (cytotoxic) T cell raised against an influenza virus peptide in an individual expressing HLA-A2 will not recognize influenza-infected HLA-A1. This concept is known as “MHC restriction” and was described by Shevach and Rosenthal for recognition of amino acid polymers,178 by Zinkernagel and Doherty for recognition of viral antigens,179,180 and by Shearer et al. for recognition of altered self-ligands.181 Given that T cells are MHC-restricted, it is difficult to understand why they should ever recognize a foreign HLA type. However, in practice they do, data indicate that such alloreactive T cells arise with remarkably high frequency. It is estimated that between 1/10 to 1/1000 activated clonally distinct T cells are capable of responding to any random allogeneic HLA molecule.182,183,184,185 Given the number of T cells in the human lymphoid system, this represents a striking tendency for T cells that are normally restricted to recognizing self-HLA molecules complexed to foreign peptides to cross-react on allogeneic HLA molecules. This cross-reaction can arise from direct recognition of the allogeneic HLA/peptide complex, which usually depends on the peptide antigen as well as the allogeneic HLA molecule. Alternatively, allorecognition by T cells can occur indirectly.186,187 In such cases, peptides derived from the allogeneic HLA molecules are presented as nominal antigen after processing by the host cells bearing self-HLA molecules. In the normal course of events, T-cell alloreactivity is an in vitro curiosity, although it is still not entirely clear why the fetal “allograft” does not stimulate the maternal immune system. However, it is the clinical transplantation of organs and hematopoietic stem cells across HLA compatibility barriers that produces graft rejection or GVHD due to T-cell alloreactivity. Fully allogeneic transplants theoretically expose the recipient immune system to up to 12 non-self-HLA allele products expressed by the allograft. Moreover, the “self-peptides” constitutively presented by allogeneic HLA molecules are likely to be quite distinct from those presented by syngeneic HLA molecules because the polymorphisms of the peptide antigen-binding cleft of the MHC-I molecule that distinguish HLA alleles alter the spectrum of selected peptides. In the presence of appropriate costimuli, allogeneic HLA/peptide complexes probably stimulate powerful T-cell responses because of the high density of unusual determinants and the diversity of new peptide ligands presented by the allogeneic HLA/peptide complexes. Because there are many MHC-linked genes encoding a host of proteins, many lacking known immunologic function, it is likely that polymorphisms in these molecules contribute significantly to the alloresponse (see previous discussion of Mhc genetic maps and Mhc alleles). Accordingly, many studies have demonstrated an incremental improvement in long-term graft survival with progressively higher levels of HLA matching at HLA-A, -B, and -DR loci. For this reason, HLA matching is essential in allogeneic HSC transplantation and highly desirable in solid-organ transplantation. The degree of HLA matching usually required for renal transplantation is shown in Figure 21.4 and for bone marrow transplantation in Table 21.5. Survival of HSC transplants varies according to the nature of the disease, disease stage, and age of the patient but can be >70% in some cases (see Centre for International Blood and Marrow Transplant Research at www.cibmtr.org/pages/index.aspx). The effect of HLA matching on solid-organ transplantation has been monitored by the Collaborative Transplant Study since 1982 (www.ctstransplant.org/).
FIG. 21.4. Effect of HLA Matching on First Deceased Donor Kidnery Graft Survival. Survival of first kidney transplants based on number of HLA-A, -B, and -C mismatches as a function of time. Courtesy of Collaborative Transplant Study, G. Opelz, University of Heidelberg (www.ctstransplant.org/public/graphics.shtml).
TABLE 21.5 MHC Matching versus Success of Bone Marrow Transplantation
MHC Compatibility
Risk of Graft Rejection (%)
Risk of Acute Graftversus-Host Disease (%)
Survival (3 Years) (%)
Share two haplotypes (HLA identical sibling)
2
40
50
Share one haplotype plus phenotypically identical
7
40
50
1 HLA mismatch
9
70
50
>2 HLA mismatches
21
80
15
Share zero haplotypes (unrelated)
“Matched”
3
80
35
“Mismatched”
5
95
35
HLA, human leukocyte antigen; MHC, major histocompatibility complex.
Reprinted from Christiansen FT, Witt CS. Allogeneic bone marrow transplantation. In: Bradley J, McCluskey J, eds. Clinical Immunology. Oxford: Oxford University Press; 1997:445, with permission.
In addition to the allogeneic cellular response, the antibody response to HLA molecules and ABO blood groups can also cause rejection of certain grafts, especially where these antibodies are preformed and, therefore, present at the time of organ transplantation. Antibodies to ABO blood-group antigens react with these determinants on vascular endothelium, and, therefore, ABO-incompatible solid organs can be rapidly rejected by humoral mechanisms. In patients who have been transfused or previously transplanted, or in multiparous females, exposure to allogeneic HLA molecules can also result in the production of anti-HLA class I antibodies. These preformed antibodies can lead to acute and hyperacute rejection of grafts expressing the particular HLA molecules recognized by these antibodies. Renal graft survival improves with fewer HLA mismatches. In Figure 21.4, cumulative data for graft survival are plotted as a function of time. Curves represent those groups with the indicated number of mismatches. Therefore, for solid-organ transplants, individuals are not only matched as closely as possible for their HLA types to avert cellular rejection but, it is also necessary to ensure ABO compatibility and to exclude preformed antidonor HLA antibodies in the host. Paradoxically, some patients who have received multiple blood transfusions prior to transplantation appear to develop some form of T-cell tolerance to allogeneic donor HLA alleles; renal graft survival is actually enhanced in these individuals. This is known as the “transfusion effect,” and in some centers, pretransplant transfusion and even donor-specific transfusions are routinely carried out.
Transfusion of potential renal transplant recipients carries the risk, however, of inducing undesirable anti-HLA antibodies in the patient. Testing for anti-HLA antibodies is known as the “crossmatch.” In practice, many laboratories crossmatch only for anti-HLA class I antibodies. Crossmatch compatibility to exclude anti-HLA class I antibodies is essential in renal transplantation and is widely practiced in heart/lung transplantation. Crossmatching for liver transplantation is practiced at only some centers, and the evidence that a positive crossmatch predicts allogeneic liver graft rejection has not convinced everyone of its importance in routine matching. Patients awaiting renal transplantation are usually monitored regularly for anti-HLA class I antibodies because the level and specificity of these antibodies can change with time. This monitoring involves regular crossmatching of patient serum against a panel of randomly selected cells bearing different HLA types. The percentage of positively reacting cells is known as the “panel reactivity.” When carrying out a crossmatch between a patient’s serum and donor cells, many centers test the current as well as “historical peak” serum from the patient. The historical peak is defined as the patient serum sample giving the highest panel reactivity throughout the monitoring period and is thought to be a reflection of previous HLA sensitization. Most centers now prescreen patients on transplant waiting lists for anti-HLA antibodies using MHC-coated beads as a source of antigen and a highly sensitive flow cytometry technology for their detection. In highly sensitized patients with multiple host antidonor antibodies, these unwanted antibodies can sometimes be functionally eliminated using B-cell ablation therapies (anti-CD20 monoclonal antibodies), plasmapharesis, and intravenous gammaglobulin infusions, thus allowing successful transplantation. Similar approaches, coupled with plasmapharesis, are now taken with increasing frequency in transplanting kidneys across the ABO barrier. The role of antibody crossmatching in allogeneic HSC transplantation is less clear, and many centers do not take the class I or class II crossmatch into account when identifying a bone marrow transplant donor. On the other hand, some large centers place considerable importance on a positive crossmatch as a predictor of bone marrow rejection, and it is therefore advisable to crossmatch bone marrow donor-recipient pairs when there is a high risk of rejection (eg, aplastic anemia). Crossmatching is also used to detect anti-HLA antibodies that may cause refractoriness to platelet transfusion with random platelets.
Family Studies in Histocompatibility Testing
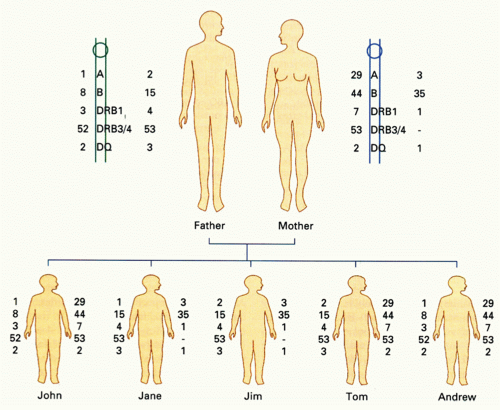
The linkage of HLA loci on chromosome 6 means that individuals will usually inherit a set of nonrecombined HLA alleles encoded at linked HLA loci from each parent. This set of genes (the haplotype) is often identifiable in family studies, where all the alleles present on one chromosome cosegregate. In identifying donors for HSC transplantation, testing of family members is essential to determine haplotypes accurately.188,189 This is because sharing of HLA antigens from different haplotypes is quite common in families, so mismatches within HLA subtypes (ie, allele-level mismatches) are easily overlooked as a result of mistaken haploidentity of siblings or other family members. Because unrecognized HLA mismatching is poorly tolerated in HSC transplantation, high-resolution sequence-based DNA matching (or its equivalent) is required to avoid GVHD.190,191,192 The impact of HLA on the outcome of HSC transplants has been studied in unrelated donor-recipient pairs by several groups,193,194 and guidelines for matching have been published by the U.S. National Marrow Donor Panel.195 These data show that unrelated matched transplants can be as effective as sibling matched donor-recipient transplants in donor survival. Mismatching beyond a single, or at most two, HLA loci is associated with increased GVHD and inversely with a lower rate of leukemic relapse. Although allogeneic HSC transplantation can be carried out across single-locus mismatches, there is little correlation with the magnitude of a given mismatch (ie, number of polymorphisms between alleles) and the subsequent immune response. Donor-recipient MHC differences of just a single amino acid can provoke strong alloresponses comparable to reactions between vastly different MHC products.191 An example of haplotyping in a family study is shown in Figure 21.5. Matching for HLA-DP in allogeneic stem cell transplantation appears to improve graft success in both stem cell and renal transplantation,196,197 but testing for this locus is not routinely carried out clinically for renal transplants and only for HSC when several donors are available and a rational choice of the best donor has to be made.198,199 HLA-C matching is also important for chronic myeloid leukemia.200 Typically, an HLA typing laboratory will test for HLA-A, -B, -DRB1, -DRB3, -DRB4, -DRB5, and -DQ loci for HSC transplant matching.198 Many centers will also insist on HLA-C testing. In the family study shown in Figure 21.5, the mother and father are mismatched at both haplotypes. Among the children, John and Andrew are haploidentical (and therefore phenotypically identical). Jane and Jim share a single haplotype, as do Tom and Jim. Jane’s paternal haplotype is a recombinant involving a crossover event between HLA-A and HLA-B. Recombination is observed between HLA-A/B and HLA-B and HLA-DR in about 1% of meiotic events. The implications of this family study are that Andrew and John would be ideal bone marrow donors for each other. However, none of the other siblings would be suitable as a donor for these brothers. Even though there is sharing of a single haplotype between Tom and both Andrew and John, the complete mismatch in the second haplotype would make Tom unsuitable as a donor for HSC transplantation, which requires very close matching of HLA. On the other hand, haplotype mismatching is common in renal transplantation, in which perfect HLA matching is not absolutely required or routinely achievable. However, for renal and other solid organ transplantation, ABO blood group compatibility is usually essential because these determinants are expressed on vascular endothelium where recognition by isohemagglutinins leads to rapid intravascular coagulation and organ failure. When a matched sibling donor does not exist for a patient requiring allogeneic HSC transplantation (70% of cases), searching of the extended family or unrelated bone marrow registries is indicated. The National Marrow Donor Panel (www.marrow.org) has several million potential donors suitable for unrelated stem cell transplantation; these donors are used in the United States and worldwide. Bone marrow donor registries also exist in Europe, Australia, Hong Kong, and Japan; these registries often provide donors for patients in other parts of the world. Mobilization of stem cells in the blood following administration of hematopoietic growth factors is now widely used to avoid the need for marrow collections from donors.
FIG. 21.5. Segregation of Haplotypes in a Family. From McCluskey,558 with permission.
In the last decade, many HSC transplant protocols have sought to reduce the pretransplant conditioning of patients. There have also been controversial reports indicating dramatic outcomes of HSC transplantation where deliberate KIR-ligand mismatching leads to donor-versus-recipient NK-cell alloreactivity toward cells of hematopoietic origin.201,202,203,204 The Velardi group in Perugia has championed this approach providing evidence that in the absence of GVHD, a high frequency of donor NK cells in KIR-ligand mismatched transplants can remove recipient’s target cells.202 In particular, HLA haplotype-mismatched hematopoietic transplants can be beneficial in adult acute myeloid leukemia and childhood acute lymphoblastic leukemia leading to the development of adoptive NK-cell transfers as an experimental therapy for graft-versus-leukemia treatment.205 However, these transplants must be rigorously T-cell depleted, and variable results appear to reflect different transplant protocols, disease parameters, and KIR-mismatching assignment.203,204,206
Cord blood banks have also been established around the world. However, cord blood donation of stem cells is often unsuitable for adult transplantation because of limitations in the volume of cord blood collections providing inadequate numbers of stem cells. Many centers will combine different cord bloods that are matched with the recipient as much as feasible to increase the stem cell number. Cord bloods offer the advantage of finding donors faster than adult unrelated registries197 and theoretically providing banked stem cells from ethnic minority groups that are not well represented in bone marrow donor registries.198 Cord blood transplants induce less GVHD than bone marrow or peripheral blood stem cell transplants, but posttransplant engraftment is slower.196,207 Cord transplants may also tolerate greater levels of HLA mismatching than unrelated bone marrow transplant, but this is dependent on the transfer of sufficient CD34+ stem cells.208 The results of transplants using bone marrow or peripheral blood stem cells are now considered comparable clinically,207 but the impact of HLA matching in patients transplanted at early stage in their disease appears to be more marked. Functional tests of HLA compatibility testing for HLA identity at all HLA loci is a daunting task for most laboratories because of the very large number of alleles present in the population. Moreover, in renal transplantation, some mismatches appear to be well tolerated and are associated with long-term graft survival, whereas other mismatches of similar genetic disparity are poorly tolerated and are associated with early rejection.209,210 Reliable methods for predicting these “taboo” mismatches are not readily available. Similarly, high-resolution HLA typing does not predict all GVHD when selecting suitable unrelated donors for HSC transplantation.190,211 Therefore, there has been a great deal of interest in developing functional or in vitro cellular tests of overall donor-recipient compatibility. Unfortunately, none of the tests so far developed provides convincing predictability of impending graft rejection or, more importantly, GVHD.
Among the tests used historically for assessing functional compatibility are the mixed lymphocyte reactivity (MLR) test and allogeneic T-helper or cytotoxic T-lymphocyte precursor studies. The MLR or MLC (C = culture) involves measuring T-cell proliferation of host T cells in response to donor lymphocytes and vice versa. In a one-way MLR, the stimulating lymphocytes are irradiated to prevent their proliferation; in the two-way MLR, both stimulator and responder cells are allowed to proliferate. In MLR studies, it is necessary to include controls showing that responder cells can all respond and that stimulator cells can all stimulate across an appropriate barrier such as third-party donor cells. Responses can vary widely, and individual laboratories use their own cutoff values to define negative (ie, nonreactive) and positive (ie, reactive) MLR results.211 Unfortunately, known HLA mismatches can be present in a negative MLR, and a positive MLR can be obtained between phenotypically HLA-identical transplant pairs. Because the MLR is biased toward measuring HLA class II discrepancies and is notoriously irreproducible,211 most laboratories have abandoned this test in favor of implementation of high-resolution polymerase chain reaction-based HLA class II typing. Measurement of allogeneic cytotoxic T-lymphocyte or helper T-cell precursor frequencies is carried out at specialized bone marrow transplant centers but is not universally accepted as being predictive of GVHD.212,213 The test is labor intensive and requires a skilled technician for reproducibility. Precursor frequencies are estimated by limiting dilution analysis or ELISpot of donor-versus-host lymphocytes (ie, T cells expected to cause GVHD). High precursor frequencies (up to 1 in 104 cells) are thought to be associated with a greater risk for acute GVHD.214 It is possible that precursor studies detect major and minor incompatibilities, and so, theoretically, they might give a broad measure of the transplant barrier, but technical improvements will be required before this test is widely adopted in clinical practice.215
The Mhc and Disease
In addition to the control of transplant acceptance or rejection and immune responsiveness, the Mhc in the human plays an important role in the etiology of a number of diseases, many of which are autoimmune in nature.123,216,217,218 Several human diseases are associated with the Mhc-III genes because some of the structural genes for enzymes involved in the adrenal steroid biosynthetic pathway (ie, 21-hydroxylase, CYP21A2) map to this region. As many as 200 diseases have well-established genetic linkages to the Mhc,217,219,220,221,222 the most important of which are summarized in Tables 21.6 and 21.7. Recent genome-wide association studies confirm the importance of the HLA-DRB1 locus in rheumatoid arthritis and type 1 diabetes,71,223,224,225 and fine mapping analysis suggests that a total of five amino acid polymorphisms (three in HLA-DRB1, one in HLA-B, and one in HLA-DPB1), all located in peptide-binding grooves, almost completely explain the MHC association with the risk of rhuematoid arthritis.226 The precise mechanisms underlying the association of most of these diseases with the particular Mhc haplotypes are unknown, but several models have been proposed, including the cross-reactivity of antimicrobial antibodies and the molecular mimicry of viral antigens that might induce T-cell responses to selfantigens.227,228,229 The very high incidence of some diseases associated with certain HLA genes assists in the diagnosis as well as the counseling of patients and their families. Several of these diseases are of particular note. Because virtually 100% of patients with narcolepsy have HLA-DQB1*06:02 (associated with HLA-DR2),230,231 HLA typing can be used as a test of disease exclusion. Thus, a diagnosis of narcolepsy can be excluded with reasonable certainty if the patient does not have HLA-DQB1*06:02. On the other hand, the presence of HLA-DQB1*06:02 is of little predictive value in diagnosis of narcolepsy because this HLA type is relatively common in many populations and occurs frequently in the absence of disease.
TABLE 21.6 Some HLA Disease Associations
Disease
MHC-I
Strength of Association
Ankylosing spondylitis
HLA-B27
+++
Reiter disease
HLA-B27
++
Psoriasis
HLA-C*06
+
Abacavir drug hypersensitivity
HLA-B*57:01
+++
Behcet disease
HLA-B*51
+
Birdshot retinopathy
HLA-A*29
+++
MHC-II
Narcolepsy
HLA-DQB1*06:02
++
Insulin-dependent diabetes mellitus
HLA-DQ8
++
HLA-DQ2
+
HLA-DR2
–
Rheumatoid arthritis
HLA-DR4
+
Celiac disease
HLA-DQ2
+++
HLA-DQ8
+
Multiple sclerosis
HLA-DR2
+
HLA, human leukocyte antigen; MHC, major histocompatibility complex
+++ = very strong association that is clinically useful as a diagnostic tool
++ = strong association with likely primary involvement in disease pathogenesis
+ = clear association with likely role in disease pathogenesis
– = negative or protective influence on disease probability
For a more detailed summary of MHC and disease associations see Shiina et al.220
TABLE 21.7 HLA Disease Associations and Relative Riska
Disease
Antigen
Estimated or Relative Risk
Graves disease or myasthenia gravis
B*08:01 DRB1*03:01 DQA1*05:01 DQB1*02:01
4 or 2.5
Multiple sclerosis
DRB1*15:01 DQB1*06:02
4
Multiple sclerosis
DQA1*01:02
4
Psoriasis
C*06:02
5
Celiac disease
DQA1*02:01/DQB* 02:02 (DQ2.2)
1
DQA1*05:01/DQB1* 02:01 (DQ2.5)
7
SLE
DRB1*15:01
2
Type 1 diabetes or SLE
DRB1*03:0101
4.5
aThese examples were taken from single nucleotide polymorphism associations reported by de Bakker et al.,71 where the tag single nucleotide polymorphisms and coefficients of determination (r2) are also reported. For celiac disease, DQ2.2 only has an effect when in combination with DQ2.5. The relative risk of DQ2.5 changes depending on the coordinate presence of other DQ types.
SLE, systemic lupus erythematosus.
Ankylosing spondylitis is so strongly associated with the Mhc-I allele HLA-B27 and the presence of some bacterial pathogens that it is a popular hypothesis that ankylosing spondylitis is due to the stimulation of particular T cells by HLA-B27-presented bacterial antigens that cross-react on self-tissues. These T cells are then thought to initiate an inflammatory cascade. Despite the strong association of HLA with spondyloarthropathy, critical evaluation of the literature brings a postinfectious etiology into question and certainly more studies are indicated.232,233 Recent genome-wide studies strongly indicate that a genetic interaction between an enzyme involved in processing MHC-I-associated peptides, ERAP1, and HLA-B27 contributes to disease susceptibility.234 The tendency of HLA-B27 to form disulfide-linked covalent dimers raises the question whether the resulting cellular pathology related to poor cell surface expression of this MHC-I molecule may be related to the inflammation of joints that is characteristic of this disease.235,236
Hereditary hemochromatosis is one of the most common genetic disorders in Caucasian populations (with a prevalence of 1/300 to 1/400). The gene controlling this condition (HFE) is Mhc linked, mapping approximately three Mb telomeric to the HLA-A locus.122 The HFE protein is a class I-like molecule, the structure of which has been determined.237,238 HFE assembles with β2m and is expressed in the intestinal mucosa and placenta, where it plays a role in regulating iron uptake and transport.239,240 Mice that are homozygous for an induced defect of β2m as well as those with targeted inactivation of the HFE gene suffer from iron overload or hemochromatosis.241,242,243,244 Mutations at loci other than β2m or HFE also lead to the same disease phenotype.245 HFE regulates the affinity of the receptor for transferrin, resulting in an alteration of the efficiency of iron transport. The most common molecular defect associated with hereditary hemochromatosis involves a point mutation that results in a Cys282Tyr substitution in the α3 domain of this class I-like molecule.122 This mutation accounts for >80% of patients with hereditary hemochromatosis.246 The disruption of the disulfide bond in the α3 domain at this site prevents efficient folding of the molecule and impairs assembly with β2m, resulting in improper HFE protein expression. This leads to a failure to downregulate the affinity of the transferrin receptor for its ligand, transferrin, presumably causing increased iron uptake by cells and tissue damage as a result of iron overload. A second common HFE mutation, 187G, results in a His63Asp substitution and a very slight increase in susceptibility to developing hereditary hemochromatosis, depending on the genotype of the individual.247 Incomplete penetrance of even the high-risk Cys282Tyr HFE genotypes can be partly explained by natural iron deficiency from limited dietary intake and menstrual losses in women.
Only gold members can continue reading. Log In or Register to continue