activity, and some miRNAs promote epigenetic methylation of cell cycle genes in mammalian cell lines.11 MicroRNAs also bind directly to DNA within promoter regions, thereby blocking transcription.12
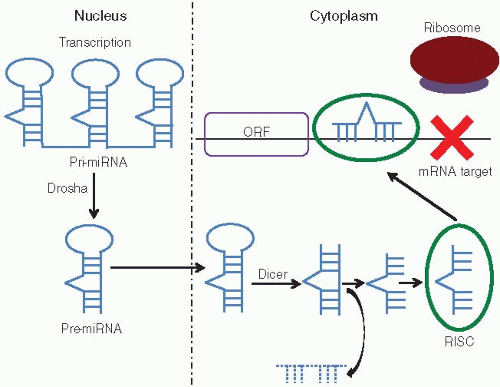 FIGURE 13.1: Primary microRNAs (pri-miRNAs) are transcribed as multimers composed of several hairpin loop structures with imperfect base pairing. These multimers are subsequently cleaved by the Drosha enzyme complex into single hairpin loops and exported to the cytoplasm. The Dicer enzyme complex processes the transcripts into asymmetric duplexes. One strand of nucleic acids is degraded and the other is loaded into the RNA-induced silencing complex (RISC), which represents the functional unit of miRNA-driven translation regulation. Each miRNA-RISC complex can bind to several different mRNA targets by sequence complementarity, thereby preventing ribosomal interaction with the open reading frame (ORF). |
of this pathway that promote constitutive activation of Wnt-mediated signaling.18 In the absence of functional APC, Wnt signaling leads to accumulation of β-catenin in the cytoplasm, which is translocated to the nucleus where it facilitates transcription of protooncogenes, such as CMYC and cyclin D1.18 Biallelic APC inactivation usually results from mutations, but may also occur due to downregulation by miRNAs. For example, both miR-135a and miR-135b are complementary
to the 3’ UTR of APC mRNA. Colon cancers with high miR-135a and miR-135b levels also contain fewer APC mRNA transcripts, suggesting that miR-135a and miR-135b regulate Wnt signaling by promoting decay of APC mRNA.19
Table 13.1 Summary of the Literature Regarding Dysregulation of MicroRNAs in Colon Cancer | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Table 13.2 Regulatory MicroRNA Targets and Function in Colon Cancer | ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Related posts:
 Embryology, Anatomy, and Normal Histology of the Colorectum and Appendix
Embryology, Anatomy, and Normal Histology of the Colorectum and Appendix
 Handling of Colorectal Cancer Resection Specimens
Handling of Colorectal Cancer Resection Specimens
 Morphologic Classification of Colorectal Epithelial Tumors
Morphologic Classification of Colorectal Epithelial Tumors
 Noninvasive Biomarkers and Early Detection of Colorectal Cancer
Noninvasive Biomarkers and Early Detection of Colorectal Cancer
 Emerging Endoscopic Techniques in the Management of IBD-related Neoplasia
Emerging Endoscopic Techniques in the Management of IBD-related Neoplasia
 Nonmucinous Epithelial Tumors of the Appendix
Nonmucinous Epithelial Tumors of the Appendix
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


