Macrophages (Mø) represent a family of mononuclear leukocytes that are widely distributed throughout the body within and outside of lymphohemopoietic organs. They vary considerably in lifespan and phenotype, depending on their origin and local microenvironment. Mature Mø are highly phagocytic, relatively long-lived cells that are adaptable in their biosynthetic responses to antigens and microbial stimuli. The functions of Mø within tissues are homeostatic, regulating the local and systemic milieu through diverse plasma membrane receptors and varied secretory products. They react to, and themselves generate, signals that influence growth, differentiation, and death of other cells, recognizing and engulfing senescent and abnormal cells. These activities contribute substantially to recognition and defense functions against invading microorganisms, foreign particulates, and other immunogens. Innate immune functions of Mø complement their contributions to acquired humoral and cellular immunity, in which they regulate activation of T-and B-lymphocytes; this is achieved in part through their specialized derivatives, dendritic cells (DCs) of myeloid origin. Mø, with or without DCs, process and present antigen; produce chemokines and cytokines such as interleukin (IL)-1, IL-6, IL-12, IL-18, IL-23, tumor necrosis factor (TNF)-&agr;, IL-10, and TGF&bgr;; and phagocytose apoptotic and necrotic cells. Acting directly or under the influence of other immune cells, Mø capture extra- and intracellular pathogens, eliminate invaders, and deliver them to appropriate subcompartments of lymphoid organs. As key regulators of the specific as well as the natural immune response, Mø boost as well as limit induction and effector mechanisms of the specific immune response by positive and negative feedback.
The properties and roles of DCs, especially in antigen presentation, are described in detail elsewhere in this volume. Here, we focus on other members of the Mø lineage, consider their interrelationship, and outline specialized properties that underlie their roles in the execution and regulation of immune responses. A number of texts and presentations deal with the history and broad aspects of Mø immunobiology.1,2,3,4,5,6,7,8
SOME LANDMARKS IN THE STUDY OF MACROPHAGES
Our understanding of Mø developed in parallel with the growth of immunology as an experimental science. Metchnikoff, a comparative developmental zoologist, is widely credited for his recognition of phagocytosis and leukocyte recruitment as a fundamental host defense mechanisms of primitive, as well as highly developed multicellular organisms.3,4,7 The Nobel awards of 2011 to Bruce Beutler,9 Jules Hoffman,10 and Ralph Steinman11 reflect the paradigm shift of immune recognition from lymphocytes to innate antigen-presenting cells (APCs). Metchnikoff already clearly stated the link between capture of infectious microorganisms by the spleen and subsequent appearance of reactive substances (antibodies) in the blood, although mistakenly ascribing their production to the phagocytes themselves. The importance of systemic clearance of particles by Mø, especially Kupffer cells in liver and other endothelial cells, was enshrined in the term reticuloendothelial system. Although it was rejected by influential investigators in the field in favor of the term mononuclear phagocyte system, the appreciation that sinuslining Mø in liver and elsewhere share common properties with selected endothelial cells is worth preserving. 3 Earlier studies by Florey and his students, including Gowans, established that circulating monocytes give rise to tissue Mø. Van Furth and his colleagues investigated the life history of Mø by kinetic labeling methods; subsequently, the development of membrane antigen markers facilitated a more precise definition of specialized Mø subpopulations in tissues such as brain. The appearance and potential importance of Mø during development also became evident as a result of sensitive immunocytochemical methods. Morphologic and functional studies by Humphrey and many others drew attention to striking diversity among Mø-like cells in secondary lymphoid organs, especially within the marginal zone of the spleen, where complex particulates and polysaccharides are captured from the circulation.
The era of modern cell biology impinged on Mø studies following the studies of Cohn,12 Hirsch, and their colleagues. Their work touched on many aspects of cell structure and function, including phagocytosis (the zipper mechanism of Silverstein), fluid- and receptor-mediated endocytosis, secretion, and antimicrobial resistance. Isolation and in vitro culture systems became available for cells from mice and humans, especially after the identification of specific growth and differentiation factors such as colony-stimulating factor-1 (CSF-1). It is perhaps fitting that the earliest known natural knockout (ko) affecting Mø, a natural mutation in the op gene in the osteopetrotic mouse, should involve CSF-1.13 Cell lines retaining some but not all features of mature Mø have been useful for many biochemical and cellular studies. Macrophages and dendritic cells can be derived from embryonic stem cells and induced pluripotent cells by growth in appropriate culture conditions and transfection of selected transcription factors.
The role of Mø as antigen-processing cells able to initiate adaptive immune responses had false trails (“immunogenic ribonucleic acid [RNA]” was thought to be involved at one time) and encompassed early genetic strategies (Mø of mice selected for high antisheep erythrocyte antibody responses by Biozzi and colleagues displayed enhanced degradative properties; adherent cells from defined guinea pig strains were shown to play an important role in major histocompatibility complex [MHC] Ia-restricted antiinsulin responses). For many years, the APC functions of adherent cells were highly controversial as promoted by Unanue, who concentrated on intracellular processing by Mø, and Steinman, who discovered the specialized role of “DCs” in antigen presentation to naive T-lymphocytes. The importance of Mø as effector cells in immunity to intracellular pathogens such as Mycobacterium tuberculosis was recognized early by Lurie and Dannenberg. Mackaness used Listeria monocytogenes and bacille Calmette-Guérin (BCG) infection in experimental models and developed the concept of Mø activation as an antigen-dependent but immunologically nonspecific enhancement of antimicrobial resistance. The subsequent delineation of T-lymphocyte subsets and characterization of interferon (IFN)-&ggr; as the major cytokine involved in macrophage activation, including MHC II induction, merged with increasing knowledge of the role of reactive oxygen and, later, nitrogen metabolites as cytotoxic agents. The role of virus-infected Mø as MHC I-restricted targets for antigen-specific CD8+ killer cells was part of the initial characterization of this phenomenon by Zinkernagel and Doherty. D’Arcy Hart was an early investigator of the intracellular interactions between Mø and invaders of the vacuolar system, especially mycobacteria, which survive within Mø by inhibiting acidification and phagosomelysosome fusion, thus evading host resistance mechanisms. Mouse breeding studies by several groups defined a common genetic locus involved in resistance to BCG, Leishmania, and Salmonella organisms. The host phenotype was shown to depend on expression in Mø and, many years later, the gene (termed N-ramp for natural resistance-associated membrane protein) was identified by positional cloning by Skamene, Gros, and their colleagues. Positional cloning by Beutler and associates led to the identification of the gene responsible for lipopolysaccharide (LPS) resistance in particular mouse strains. Together with studies by Hoffmann and his colleagues on the toll pathway in Drosophila, this work resulted in an explosion of interest in the identification of mammalian tolllike receptors (TLRs) and their role in innate immunity to infection. At the same time, it became apparent that some malignant tumors contain macrophage populations that may favor their growth.
This brief survey concludes with the identification of Mø as key target cells for infection, dissemination, and persistence of human immunodeficiency virus (HIV), tropic for Mø by virtue of their expression of cluster of differentiation (CD)4, chemokine coreceptors, and DCSIGN, a C-type lectin also expressed by DCs. Although Mø had been implicated by earlier workers such as Mims as important in antiviral resistance generally, their role in this regard was neglected before the emergence of HIV as a major pathogen.
Many molecules have been identified as important in Mø functions in immunity and serve as valuable markers to study their properties in mice and humans. These include Fc and complement receptors, which are important in opsonic phagocytosis, killing, and immunoregulation; scavenger receptors originally implicated in foam cell formation and atherogenesis by Brown and Goldstein; nonopsonic lectin receptors, such as the mannose receptor (MR) and &bgr;-glucan receptor (dectin-1) and secretory products such as lysozyme, neutral proteinases, TNF&agr;, chemokines, and many other cytokines. A range of membrane antigens expressed by human and rodent mononuclear phagocytes has been characterized and reagents made available for further study of Mø in normal and diseased states. Recently, the role of deoxyribonucleic acid (DNA)-binding transcription factors including members of the NF-&kgr;B and ETS (Pu-1) families has received increased attention in the study of differential gene expression by Mø. Gene inactivation has confirmed the important role of many of these molecules within the intact host, and use has been made of cell-specific or conditional ko to uncover the role of Mø in immunologic processes. Naturally occurring inborn errors in humans such as the leukocyte adhesion deficiency syndrome and chronic granulomatous disease have contributed to the analysis of important leukocyte functions, including those of Mø, in host resistance to infection. Mutations in a monocyte-expressed gene (nucleotide oligomerization domain [NOD]-2), involved in cytosolic sensing of microbial products and NF-&kgr;B activation, have been implicated in a subset of individuals with an enhanced susceptibility to Crohn disease. The validity of murine ko models for human genetic deficiencies has been confirmed for key molecules involved in Mø activation, such as IFN&ggr; and IL-12. N-ethyl N-nitrosourea mutagenesis has begun to reveal new macrophage innate immune functions, as has increasing application of system biology tools for microarray, proteomic, epigenetic, and microRNA analysis.
PROPERTIES OF MACROPHAGES AND THEIR RELATION TO IMMUNE FUNCTIONS
Introduction
Mø participate in the production, mobilization, activation, and regulation of all immune effector cells. They interact reciprocally with other cells while their own properties are modified to perform specialized immunologic functions. As a result of cell surface and auto- and paracrine interactions, Mø display marked heterogeneity in phenotype,14,15 a source of interest and considerable confusion to the investigator. Increasing knowledge of cellular and molecular properties of Mø bears strongly on our understanding of their role in the immune response. These will be reviewed briefly, with emphasis on functional significance, and attention will be drawn to unresolved and controversial issues.
Growth and Differentiation: Life History and Turnover
In contrast to T- and B-lymphocytes, monocytes from blood give rise to terminally differentiated Mø that cannot recirculate or reinitiate DNA replication except in a limited way. Unlike other myeloid granulocytic cells, Mø can be long lived and retain the ability to synthesize RNA and protein to a marked extent, even when in a relatively quiescent state as “resident” cells. These are distributed throughout the tissues of the body and constitute a possible alarm-response system, but they also mediate homeostatic and poorly understood trophic functions. Following inflammatory and immune stimuli, many more monocytes can be recruited to local sites and give rise to “elicited” or “immunologically activated” Mø with altered surface, secretory, and cytotoxic properties. The origins of Mø from precursors are well known: from yolk sac (and possibly earlier paraaortic progenitors), migrating to fetal liver, then spleen and bone marrow, before and after birth.16 Yolk sac precursor cells may contribute to the establishment of selected tissue macrophages such as Langerhans cells in the adult.17 In the fetus, mature Mø proliferate actively during tissue remodeling in developing organs. In the normal adult, tissue Mø do not self-renew extensively except in specialized microenvironments such as epidermis,18 nervous system, or lung; after TH2-type parasitic infection, there can be considerable further replication at local sites of inflammation.19 Growth and differentiation are tightly regulated by specific growth factors and their receptors (eg, IL-3, CSF-1, granulocyte-macrophage [GM]-CSF/IL-34, IL-4, IL-13) and inhibitors (eg, IFN&agr;/&bgr;, transforming growth factor [TGF]-&bgr;, leukemia inhibitory factor), which vary considerably in their potency and selectivity. These processes are modulated by interactions with adjacent stromal and other cells (eg, through c-kit/ligand and Flt-3/ligand interactions). The growth-response of the target cell to an extrinsic stimulus decreases progressively and markedly (from 108 or more to 100) during differentiation from stem cell to committed precursor to monoblast, monocyte, and Mø, yet even the most terminally differentiated Mø such as microglial cells can be “reactivated” to a limited extent by local stimuli. Elicited/activated Mø respond more vigorously than resident Mø to growth stimuli in vivo and in vitro, but the molecular basis for their enhanced proliferation is unknown.
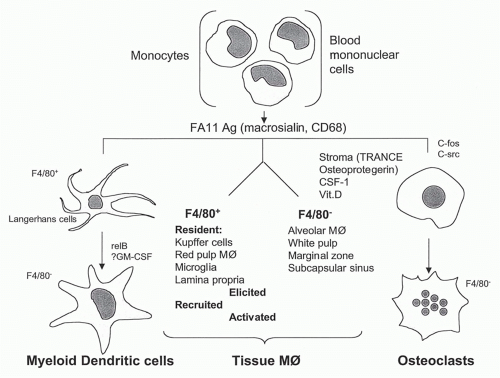
FIG. 19.1. Differentiation of mononuclear phagocytes based on antigen markers FA-11 (macrosialin, murine CD68) and F4/80. See text for detailed discussion.
Although this general picture of blood monocyte-totissue Mø differentiation has been accepted for some time as a result of parabiosis, adoptive transfer, and irradiation-reconstitution experiments, recent studies in mouse and man have demonstrated monocyte heterogeneity and distinct properties,20,21,22,23 with a subpopulation remaining within the vasculature, to perform a patrolling function. Our understanding of DCs and osteoclast differentiation is still compatible with a relatively simple model (Fig. 19.1) in which major Mø populations in mouse tissues can be characterized by selected antigen markers such as F4/80 (Emrl, a member of a family of EGF-TM7 molecules) and macrosialin (CD68), a pan-Mø endosomal glycoprotein related to the lysosome-associated membrane protein (LAMP) family. The DCs of myeloid origin (see elsewhere in this volume) share many properties with monocyte/macrophages,24 but are specialized to capture, process, and present antigens to naïve lymphocytes. Circulating precursors of DCs and and macrophages are normally present in the mononuclear fraction of blood in small numbers25; studies in the mouse may not reflect the origin and differentiation of precursor cells in humans.26 Monocytes that have crossed the endothelium may be induced to “reverse migrate” into the circulation by selected stimuli in tissues.27 Finally, the mouse spleen has been shown to serve as a reservoir of monocyte/Mø for recruitment to sites of inflammation.28
Circulating mononuclear precursors for osteoclasts are less defined and differentiate into mononucleate cells, recruited in response to sphingosine-1-phosphate to bone, for example,29 where they fuse to form multinucleate bone-resorbing osteoclasts.30 Local stromal cells, growth factors such as CSF-1, steroids (vitamin D metabolites), and hormones (eg, calcitonin, for which osteoclasts express receptors) all contribute to local maturation. Osteoprotegerin, a naturally occurring secreted protein with homology to members of the TNF-receptor family, interacts with TRANCE, a TNF-related protein, to regulate osteoclast differentiation and activation in vitro and in vivo.
Use of antigen markers such as CD34 on progenitors, CD14 and CD16 on monocytes, and chemokine receptors and multichannel fluorescein-activated cell sorter analysis have made it possible to isolate leukocyte subpopulations and study their progeny and differential responses in different mouse tissues and models of disease.31 The mononuclear fraction of blood may contain precursors of other tissue cells, including mesenchymal stem cells able to synthesize matrix proteins such as collagen, and some endothelial cells. Perhaps the mysterious follicular dendritic cells (FDCs) with mixed hemopoietic and mesenchymal properties fall in this category.
The large-scale production of immature and mature DClike cells from bulk monocytes in cytokine-supplemented culture systems (IL-4, GM-CSF, TNF&agr;) has revolutionized the study of these specialized APCs. Individually, the same cytokines give rise to Mø-like cells, and early during in vitro differentiation, the cellular phenotype is reversible. Later, when mature DCs with high MHC II, APC function, and other characteristic markers are formed, differentiation is irreversible. This process is independent of cell division, although earlier progenitors in bone marrow and GM-CSF-mobilized blood mononuclear cells can be stimulated to multiply, as well as differentiate, in vitro. These examples of terminal differentiation observed with DCs and osteoclasts may extend to other specialized, more obvious Mø-like cells. Mature Mø can be derived by growth and differentiation in steroid-supplemented media in Dexter-type long-term bone marrow cultures that contain stromal fibroblasts and hemopoietic elements. These Mø express adhesion molecules responsible for divalent cation-dependent cluster formation with erythroblasts (EbR). This receptor, possibly related to V-CAM, cannot be induced on terminally differentiated peritoneal Mø if these are placed in the same culture system. This contrasts sharply with the ready adaptation of many tissue Mø to conventional cell culture conditions, when the cells often adopt a common, standard phenotype. Irreversible stages of Mø differentiation may therefore occur in specialized microenvironments in vitro or in vivo.
Little is known about determinants of Mø longevity and turnover. Growth factors such as CSF-1 enhance Mø survival and prevent induction of an apoptotic program. The expression of Fas-L and Fas on Mø has been less studied than on lymphocytes; they and other members of the TNF and its receptor family may play a major role in determining Mø survival, especially in induced populations, where cell turnover is markedly enhanced. Tissue Mø vary greatly in their lifespan, from days to months. Apart from inflammatory and microbial stimuli, local and systemic environmental factors such as salt loading and hormones, including estrogen, are known to influence Mø turnover.
Tissue Distribution and Phenotypic Heterogeneity of Resident Macrophages in Lymphoid and Nonlymphoid Organs
The use of the F4/80 plasma membrane antigen made it possible to detect mature Mø in developing and adult murine tissues and define their anatomic relationship to other cells in endothelium, epithelium, and connective tissue, as well as the nervous system.32,33 Subsequently, other membrane antigens,34 macrosialin, sialoadhesin, and others were identified as useful markers for Mø in situ (Table 19.1). Mø subpopulations in different tissues display considerable heterogeneity in expressing these and selected receptor antigens (eg, complement receptor [CR]3 and class A scavenger receptor [SR-A]), drawing attention to unknown mechanisms of homing, emigration, and local adaptation to particular microenvironments. From the viewpoint of immune responses, a few aspects deserve comment.
Fetal Liver and Bone Marrow
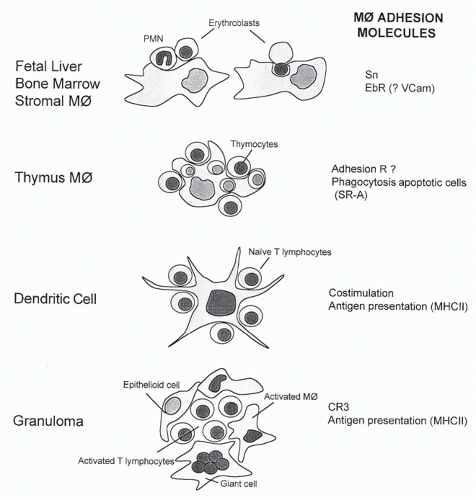
Mature Mø form an integral part of the hemopoietic microenvironment and play a key role in the production, differentiation, and destruction of all hemopoietic cells. The fetal liver is a major site of definitive erythropoiesis from midgestation.16 The bone marrow becomes active in the production of hemopoietic cells from shortly before birth, and Mø are a prominent component of the hemopoietic stroma throughout adult life. Mature “stromal” Mø in fetal liver and adult bone marrow express nonphagocytic adhesion molecules such as sialoadhesin (Sn), an immunoglobulin (Ig)-superfamily sialic acid-binding lectin (Table 19.1), and the EbR referred to previously, which is also involved in adhesion of developing myeloid and possibly lymphoid cells (Fig. 19.2). VLA-4 has been implicated as a ligand for EbR. Ligands for Sn include CD43 on developing granulocytes and on lymphocyte subpopulations. Sn clusters at sites of contact between stromal Mø and myeloid but not erythroid cells. Chemokines are able to induce polarized expression of adhesion molecules such as intercellular adhesion molecules and CD43 in leukocytes, but the significance of altered ligand distribution for interactions between Mø and bound hemopoietic cells is unknown. Adhesion of immature cells to stromal Mø may play a role in regulating their intermediate stages of development before release into the bloodstream, whereas fibroblasts in the stroma associate with earlier progenitors, as well as with Mø. Discarded nuclei of mammalian erythroid cells are rapidly engulfed by stromal Mø, but the receptors involved in their binding and phagocytosis are unknown. Mø also phagocytose apoptotic hemopoietic cells generated in bone marrow, including large numbers of myeloid and B cells. We know little about the plasma membrane molecules and cytokine signals operating within this complex milieu, but it is clear that stromal Mø constitute a neglected constituent within the hemopoietic microenvironment.
TABLE 19.1 Selected Differentiation Antigens Used to Study Murine Macrophage Heterogeneity
Ab
Ag
Structure
Ligands
Cellular Expression
Function
Comment
F4/80
F4/80 (EMR1)
EGF-TM7
?
Mature Mø, absent T areas
Peripheral tolerance
Useful marker development, CNS
FA-11
Macrosialin (CD68)
Mucin-LAMP
OX-LDL
Pan-Mø, DC
Late endosomal
Glycoforms regulated by inflammation and phagocytosis
5C6
CR3 (CD11b, CD18)
&bgr;2-integrin
iC3b, ICAM
Monocytes, microglia, PMN, NK cells
Phagocytosis, adhesion
Important in inflammatory recruitment, PMN apoptosis
2F8
SR-A (I, II)
Collagenous, type II glycoprotein
Polyanions, LTA, LPS, bacterial proteins
Adhesion, endocytosis
Protects host against LPS-induced shock
Isoforms differ, cysteine-rich domain
Modified proteins &bgr;-amyloid apolipoprotein A, E
Mø, sinusoidal endothelium
Phagocytosis of apoptotic cells and bacteria
Promotes atherosclerosis
SER-4
Sn (Siglec-1)
Ig superfamily
Sialyl glycoconjugates (eg, CD43)
Subsets tissue Mø
Lectin
Strongly expressed
3D6
Marginal zone metallophils in spleen and subcapsular sinus of lymph nodes
FIG. 19.2. Associations of tissue macrophages with other hemopoietic cells to illustrate variations on a common theme. See text for details.
Thymus
Apart from their remarkable capacity to remove apoptotic thymocytes, the possible role of Mø in positive and negative selection of thymocytes has been almost totally overlooked; more attention has been given to local DCs and their specialized properties. Mature Mø with unusual features are also present in cortex and medulla. Clusters of viable thymocytes and Mø can be isolated from the thymus of young animals by collagenase digestion and adherence to a substratum (see Fig. 19.2). The nonphagocytic adhesion receptors responsible for cluster formation are more highly expressed by thymic than other Mø, but their nature is unknown (N. Platt, unpublished observations). These Mø also express MHC class II antigens and other receptors such as the SR-A (see subsequent discussion), which contributes to phagocytosis of apoptotic thymocytes in vitro, but is redundant in vivo; other markers, such as the F4/80 antigen, are poorly expressed in situ but can be readily detected after cell isolation. A striking difference between thymic and several other tissue Mø subpopulations is their independence of CSF-1; the CSF-1-deficient op/op mouse lacks osteoclasts and some Mø populations, including monocytes, peritoneal cells, and Kupffer cells, but contains normal numbers of thymic Mø, as well as DCs and selected Mø in other sites. A second ligand for the CSF-1 receptor Fms, IL-34, may account for CSF-1 independence.35 Factors involved in constitutive recruitment of thymic Mø are unknown; following death of thymocytes induced by ionizing radiation or glucocorticoids, intensely phagocytic Mø appear in large numbers; it is not known what proportion arises locally and by recruitment.
Spleen
From the viewpoint of the Mø, the spleen is perhaps the most complex organ in the body.36,37 It contributes to hemopoiesis, which persists postnatally in some species or can be induced by increased demand, can serve as a reservoir as noted previously, and contributes to the turnover of all blood elements at the end of their natural lifespan. The spleen filters a substantial proportion of total cardiac output, captures particulate and other antigenic materials from the bloodstream, and plays an important role in natural and acquired humoral and cellular immunity. The organ is rich in subpopulations of Mø that differ in microanatomic localization, phenotype, life history, and functions (Fig. 19.3). Mø are central to antigen capture, degradation, transport, and presentation to T- and B-lymphocytes, and contribute substantially to antimicrobial resistance. Recent work has unveiled an unexpected role in facilitating activation of other lymphocyte subsets, such as invariant natural killer T cells37; CD 169+ macrophages also activate CD8 T cells in response to dead cell-associated antigens in lymph nodes and by transferring antigen to DCs in the spleen. Because other hemopoietic and secondary lymphoid organs can replace many of these functions after maturation of the immune system, the unique properties of the spleen have been mainly recognized in the immature host and in immune responses to complex polysaccharides. Splenectomy in the adult renders the host susceptible to infection by pathogenic bacteria such as pneumococci that contain saccharide-rich capsular antigens; the marginal zone of the spleen in particular may play an essential role in this aspect of host resistance.
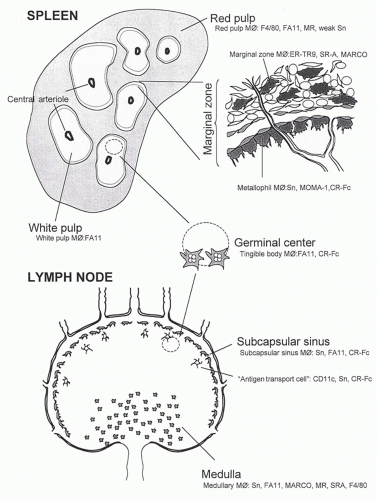
The properties of Mø in the unstimulated mature mouse spleen are different according to their localization in red or white pulp and the marginal zone. Mø are intimately associated with the specialized vasculature. Species differences in splenic anatomy and phenotype are well recognized, although Mø display broadly common features in humans and rodents. Subpopulations of Mø, DCs, and cells with mixed phenotypes have been characterized by in situ analysis by antigen markers, liposome or diphtheria toxin-depletion studies, various immunization and infection protocols, and cytokine and receptor gene ko models in the mouse. The results raise questions about the dynamics and molecular basis of cell production, recruitment, differentiation, emigration, and death within each distinct splenic compartment. Cell isolation methods are still primitive in correlating in vitro properties with those of Mø subpopulations in vivo and remain an important challenge. Detailed aspects of splenic architecture, DC origin and function, and T- and B-lymphocyte induction and differentiation are described elsewhere in this volume. Here, some features of Mø in the normal and immunoreactive organ are highlighted.
Marginal Zone Macrophages
The marginal zone of spleen consists of a complex mixture of resident cells (reticular and other fibroblasts, endothelium), Mø, DCs, and lymphoid cells, including subpopulations of B-lymphocytes. It constitutes an important interface with the circulation that delivers cells, particulates, or soluble molecules directly into the marginal sinus or via the red pulp. Resident Mø are present as specialized metallophilic cells in the inner marginal zone, and other Mø are found in the outer zone; the latter may be more phagocytic. Sn is strongly expressed by the marginal metallophils, compared with only weak expression in red pulp and virtual absence in the white pulp. Sn+ cells appear in this zone 2 to 4 weeks postnatally in the mouse as the white pulp forms. Liposomes containing clodronate, a cytotoxic drug, can be delivered systemically and deplete Sn+ cells and other Mø; regeneration of different Mø subpopulations in spleen occurs at different times, and this procedure has been used to correlate their reappearance with distinct immunologic functions. Marginal zone Mø lack F4/80 but may express an undefined ligand for F4/80 on circulating activated DCs, which mediates peripheral tolerance to anterior chamber or gut-derived antigens.38 Marginal zone Mø express phagocytic receptors, such as SR-A, which is more widely present on tissue Mø, as well as MARCO, a distinct collagenous scavenger receptor, which is almost exclusively present on these Mø in the normal mouse. The structures and possible role of these pattern recognition receptors in uptake of microbes are discussed subsequently. In vivo studies have shown that an Mø lectin, the MR, may be involved in transfer of mannosylated ligands to the site of an immune response in the white pulp.39 The MR contains a highly conserved cysteine-rich domain, not involved in mannosyl recognition, that reacts strongly with ligands on a subset of marginal metallophilic Mø, sulfated glycoforms of Sn, and CD45, among others; this has been demonstrated with a chimeric probe of the cysteine-rich domain of the MR and human Fc (CR-Fc) and by immunochemical analysis of tissue sections and affinity chromatography of spleen ligands. After immunization, this probe additionally labels undefined cells in the FDC network of germinal centers, as well as tingible body Mø. It is possible that marginal zone Mø can be induced to migrate into white pulp as described after LPS injection; alternatively, they may shed complexes of soluble MR-glycoprotein ligand for transfer to other CR-Fc+ cells, which may be resident or newly recruited mononuclear cells. Finally, the marginal metallophilic Mø population depends on CSF-1 for its appearance and on members of the TNF receptor family, as shown with op/op and experimentally produced ko mice.
FIG. 19.3. Microheterogeneity of macrophages in spleen, resting, and antigen-stimulated lymph nodes. See text for markers and details.
White Pulp Macrophages
The F4/80 antigen is strikingly absent on murine white pulp Mø, which do express FA-11 (macrosialin), the murine homolog of CD68. Actively phagocytic Mø express this intracellular glycoprotein in abundance compared with DCs. After uptake of a foreign particle (eg, sheep erythrocytes or an infectious agent, such as BCG or Plasmodium yoellii), white pulp Mø become more prominent, although it is not known whether there is migration of cells into the white pulp or transfer of phagocytosed material and reactivation of previous resident Mø. Tingible body Mø appear to be involved in uptake and digestion of apoptotic B-lymphocytes.
Red Pulp Macrophages
These express F4/80 antigen and MR strongly and in the mouse include stromal-type Mø involved in hemopoiesis. Extensive phagocytosis of senescent erythrocytes results in accumulation of bile pigments and ferritin, and play an important role in iron turnover40 and tolerance.41 The role of various phagocytic receptors in clearance of host cells and pathogens by red pulp Mø requires further study.
There is no evidence that Mø, other than interdigitating DCs, associate directly with CD4+ T-lymphocytes in the normal spleen. Following infection by BCG, for example, or by other microorganisms such as Salmonella, there is massive recruitment and local production of Mø, many of which associate with T-lymphocytes. Newly formed granulomata often appear first in the marginal zone (focal accumulations of activated Mø and activated T cells). As infections spread into the white and red pulp, the granulomata become confluent and less localized, obscuring and/or disrupting the underlying architecture of the spleen. The possible role of activated Mø in T-cell apoptosis and clearance in spleen has not been defined.
Lymph Nodes
F4/80 antigen is relatively poorly expressed in lymph node (see Fig. 19.3), but many macrosialin (CD68)+ cells are present. The subcapsular sinus is analogous to the marginal zone and contains strongly Sn+ cells; this is the site where afferent lymph enters, containing antigen and migrating DCs derived from skin and mucosal surfaces. The medulla contains Sn+, CD68+ Mø, which also express high levels of SR-A. As in the spleen marginal zone, subcapsular sinus Mø are strongly labeled by the CR-Fc probe. Following primary or secondary immunization, the staining pattern moves deeper into the cortex and eventually becomes concentrated in germinal centers. The kinetics of this process strongly suggests a transport process by Mø-related cells resembling antigen transport cells described previously. CR-Fc+ cells can be isolated by digestion of lymph nodes and form clusters with CR-Fc- lymphocytes. Adoptive transfer has shown that fluorescein-activated cell sorter-isolated CR-Fc+ cells resemble DCs in their ability to home to T-cell areas and to present antigen to naive T and B cells. Overall, there is considerable heterogeneity in the population of migratory APCs involved in antigen capture, transport, and delivery to T and B cells, and it may turn out that specialized tissue Mø as well as myeloid-type DCs can migrate in response to immunologic stimuli, especially TLR ligands.42
Peyer Patch
Although less studied, the Mø in Peyer patch resemble the CD68+, F4/80- cells described in spleen and white pulp and in other T-cell-rich areas. They are well placed to interact with gut-derived antigens and pathogens taken up via specialized epithelial M cells in the dome, and deliver antigens to afferent lymphatics, as myeloid DCs. These cells are distinct from abundant F4/80+ cells in the lamina propria found all the way down the gastrointestinal tract and may play a role in the induction of mucosal immunity. Recent studies have described heterogeneous populations of resident and recruited macrophages and DC in the mouse intestine.43 The role of the microbiome44 has received a great deal of attention in regard to innate cell phenotype and epithelial integrity in the gut.
Nonlymphoid Organs
Regional F4/80+ and CD68+ Mø are well described in liver (Kupffer cells), dermis, neuroendocrine and reproductive organs, and serosal cavities, where they are able to react to systemic and local stimuli. In the lung, alveolar Mø are strongly CD68+ but only weakly F4/80+ and are distinct from interstitial Mø and intraepithelial DCs. In the lamina propria of the intestine, Mø display a downregulated phenotype, ascribed to TGF&bgr; of local origin.45 In addition, resident Mø are found throughout connective tissue and within the interstitium of organs, including heart, kidney, and pancreas. These cells vary greatly depending on their local microenvironment; for example, in the central nervous system, microglia within the neuropil differ strikingly from Mø in the meninges or choroid plexus.46 Perivascular Mø in the brain can be distinguished from resident microglia by their expression of endocytic receptors (eg, the SR-A and MR, and of MHC I and II antigens). Microglia are highly ramified, terminally differentiated cells of monocytic origin; many Mø markers are downregulated. Their phenotype is influenced by the blood-brain barrier, normally absent in circumventricular organs, and disrupted by inflammatory stimuli. Microglia can be reactivated by local LPS and neurocytotoxins; they are then difficult to distinguish from newly recruited monocytes, which acquire microglial features once they enter the parenchyma of the brain. Resting microglia are unusual among many tissue Mø in that they constitutively express high levels of CR3 and respond to CR3 ligands, such as mAb, by induced DNA synthesis and apoptosis. In other sites, such as lung and liver, CR3 expression is a feature of recent myeloid recruitment, including monocytes. Resident Kupffer cells lack constitutive CR3 but express a novel CR implicated in clearance function.
Resident tissue macrophages in human tissues express CD68 antigen, but their phenotypic diversity and microheterogeneity in different organs remain poorly defined. Access to skin biopsies, bronchoalveolar lavage, and placenta, for example, provides material for further analysis.
Enhanced Recruitment of Monocytes by Inflammatory and Immune Stimuli: Activation in Vivo
In response to local tissue and vascular changes, partly induced by resident Mø during (re)activation by inflammatory, infectious, and immunologic stimuli, monocytes are recruited from marrow pools and blood in increased numbers; they diapedese and differentiate into Mø with altered effector functions as they enter the tissues.47 These Mø are classified as “elicited” when cells are generated in the absence of IFN&ggr; and as “immunologically activated” after exposure to IFN&ggr;. Enhanced recruitment can also involve that of other myeloid or lymphoid cells; selectivity of the cellular response depends on the nature of the evoking stimulus (immunogenic or not), the chemokines produced, and the receptors expressed by different leukocytes. Mø and other cells produce a range of different chemokines and express multiple seven-transmembrane, G protein-coupled chemokine receptors. The chemokines can also act in the marrow compartment, especially if anchored to matrix and glycosaminoglycans, may display other growth regulatory functions, and can control egress. Locally bound or soluble chemokines induce the surface expression and activity of adhesion molecules on circulating white cells, as well as directing their migration through and beyond endothelium. Feedback mechanisms from periphery to central stores and within the marrow stroma may depend on cytokines and growth factors such as macrophage inflammatory protein-1&agr; and GM-CSF, which inhibit or enhance monocyte production, respectively. The adhesion molecules involved in recruitment of monocytes, originally defined by studies in humans with inborn errors and by use of inhibitory antibodies in experimental animal models,48 overlap with those of polymorphonuclear neutrophils and lymphocytes and include L-selectin, &bgr;2-integrins, especially CR3, CD31, an Ig-superfamily molecule, and CD99; additional monocyte adhesion molecules for activated endothelium include CD44, vascular cell adhesion molecules, &bgr;1-integrins, and newly described receptors such as EMR2 and CD97, members of the EGF-TM7 family.33 The mechanisms of constitutive entry of monocytes into developing and adult tissues, in the absence of an inflammatory stimulus, are unknown.
Only gold members can continue reading. Log In or Register to continue