In 2001, the first tyrosine-kinase inhibitor imatinib was approved for clinical use in chronic myeloid leukemia. The spectacular success of this first-in-class agent ushered in a transformation in cancer drug discovery from efforts that were largely based on novel cytotoxic chemotherapy agents to an almost exclusive focus on molecularly targeted agents across the pharmaceutical and biotechnology industry and academia. This chapter summarizes this remarkable progress in this field over ˜15 years, with the focus on the concepts underlying this paradigm shift as well as the considerable challenges that remain (Table 22.1). Readers in search of more specific details on individual drugs and their indications should consult the relevant disease-specific chapters elsewhere in this volume as well as references cited within this chapter. Readers should also note that the epidermal growth factor receptor (EGFR) and human epidermal growth factor receptor 2 (HER2) receptor tyrosine kinases covered here have also been successfully targeted by monoclonal antibodies that engage these proteins at the cell surface. These drugs, referred to as biologics rather than small molecule inhibitors, are covered in other chapters. The chapter is organized around kinase targets rather than diseases and, intentionally, has a historical flow to make certain thematic points and to illustrate the broad lessons that have been and continue to be learned through the clinical development of these exciting agents.
Perhaps the most stunning discovery from the clinical trials of the Abelson murine leukemia (ABL) kinase inhibitor imatinib was the recognition that tumor cells acquire exquisite dependence on the breakpoint cluster region protein BCR-ABL fusion oncogene, created by the Philadelphia chromosome translocation.1 Although this may seem intuitive at first glance, consider the fact that the translocation arises in an otherwise normal hematopoietic stem cell, the survival of which is regulated by a complex array of growth factors and interactions with the bone marrow microenvironment. Although BCR-ABL clearly gives this cell a growth advantage that, over years, results in the clinical phenotype of chronic myeloid leukemia, there was no reason to expect that these cells would depend on BCR-ABL for their survival when confronted with an inhibitor. In the absence of BCR-ABL, these tumor cells could presumably rely on the marrow microenvironment, just like their normal, nontransformed neighbors. Thus, it seemed more likely that, by shutting down the driver oncogene, BCR-ABL inhibitors might halt the progression of chronic myeloid leukemia but not eliminate the preexisting tumor cells. In fact, chronic myeloid leukemia (CML) progenitors are eliminated after just a few months of anti-BCR-ABL therapy, indicating they are dependent on the driver oncogene for their survival and have “forgotten” how to return to normal. This phenomenon, subsequently documented in a variety of human malignancies, is colloquially termed oncogene addiction.2 Although the molecular basis for this addiction still remains to be defined, the notion of finding an Achilles’ heel for each cancer continues to captivate the cancer research community and has spawned a broad array of efforts to elucidate the molecular identity of these targets and discover relevant inhibitors.
EARLY SUCCESSES: TARGETING CANCERS WITH WELL-KNOWN KINASE MUTATIONS (BCR-ABL, KIT, HER2)
From the beginning, clinical trials of imatinib were restricted to patients with Philadelphia chromosome-positive chronic myeloid leukemia. For what seem like obvious reasons, there was never any serious discussion about treating patients with Philadelphia chromosome-negative leukemia because the assumption was that only patients with the BCR-ABL fusion gene would have a chance of responding. This was clearly a wise decision because hematologic response rates approached 90% and cytogenetic remissions were seen in nearly half of the patients in the early phase studies.3 It was obvious that the drug worked, and imatinib was approved in record time. Unwittingly, the power of genome-based patient selection was demonstrated in the clinical development of the very first kinase inhibitor. As we will see, it took nearly a decade for this lesson to be fully learned. Today, the much larger clinical experience, with an array of different kinase inhibitors across many tumor types, has led to a much better understanding of the principles that dictate oncogene addiction that, in retrospect, were staring us in the face. Foremost among them is the notion that tumors with a somatic mutation or amplification of a kinase drug target are much more likely to be dependent on that target for survival. Hence, a patient whose tumor has such a mutation is much more likely to respond to treatment with the appropriate inhibitor. This has also led to a new paradigm at the regulatory level of drug approval requiring codevelopment of a companion diagnostic (a molecularly based diagnostic test that reliably identifies patients with the mutation) with the new drug.
After chronic myeloid leukemia, the next example to illustrate this principle was gastrointestinal stromal tumor (GIST), which is associated with mutations in the KIT tyrosine-kinase receptor or, more rarely, in the platelet-derived growth factor (PDGF) receptor.4,5 Serendipitously, imatinib inhibits both KIT and the PDGF receptor; therefore, the clinical test of KIT inhibition in GIST followed quickly on the heels of the success in CML.6 In retrospect, the rapid progress made in these two diseases was based, in part, on the fact that the driver molecular lesion (BCR-ABL or KIT mutation, respectively) is present in nearly all patients who are diagnosed with these two diseases. The molecular analysis merely confirmed the diagnosis that was made using standard clinical and histologic criteria. Consequently, clinicians could identify the patients most likely to respond based on clinical criteria rather than rely on an elaborate molecular profiling infrastructure to prescreen patients. Consequently, clinical trials evaluating kinase inhibitors in CML and GIST accrued quickly, and the therapeutic benefit became clear almost immediately.
The notion that molecular alteration of a driver kinase determines sensitivity to a cognate kinase inhibitor was further validated during the development of the dual EGFR/HER2 kinase inhibitor lapatinib. Clinical trials of this kinase inhibitor were conducted in women with advanced HER2-positive breast cancer based on earlier success in these same patients with the monoclonal antibody trastuzumab, which targets the extracellular domain of the HER2 kinase. Lapatinib was initially approved in combination with the cytotoxic agent capecitabine for women with resistance to trastuzumab,7 and then was subsequently approved for frontline use in metastatic breast cancer in combination with chemotherapy or hormonal therapy, depending on estrogen receptor status. A key ingredient that enabled the clinical development of lapatinib was the routine use of HER2 gene amplification testing in the diagnosis of breast cancer, pioneered during the development of trastuzumab several years earlier. This widespread clinical practice allowed for the rapid identification of those patients most likely to benefit. If lapatinib trials had been conducted in unselected patients, the clinical signal in breast cancer would likely have been missed.
TABLE 22.1 Kinase Inhibitors: Approved or Anticipated Approval In 2014
Target
Drug
Approved Indications
Anticipated Future Indications
ALK
Crizotinib
Ceritinib
ALK mutant lung cancer
ALK mutant neuroblastoma, anaplastic lymphoma
BCR-ABL
Imatinib
Dasatinib
Nilotinib
Bosutinib
Ponatinib
Chronic myeloid leukemia
Philadelphia chromosome-positive acute lymphoid leukemia
aApproval is anticipated based on positive phase 3 data and announcement of accepted Food and Drug Administration submission by the sponsor.
The Serendipity of Unexpected Clinical Responses: EGFR in Lung Cancer
In contrast to the logical development of imatinib and lapatinib in molecularly defined patient populations, the EGFR kinase inhibitors gefitinib and erlotinib entered the clinic without the benefit of such a focused clinical development plan. Although considerable preclinical data implicated EGFR as a cancer drug target, there was little insight into which patients were most likely to benefit. The first clue that EGFR inhibitors would have a role in lung cancer came from the recognition by several astute clinicians of remarkable responses in a small fraction of patients with lung adenocarcinoma.8 Further studies revealed the curious clinical circumstance that those patients most likely to benefit tended to be those who never smoked, women, and those of Asian ethnicity.9 Clearly, there was a strong clinical signal in a subgroup of patients, who could perhaps be enriched based on these clinical features, but it seemed that a unifying molecular lesion must be present. Three academic groups simultaneously converged on the answer. Mutations in the EGFR gene were detected in the 10% to 15% of patients with lung adenocarcinoma who had radiographic responses.10,11,12 It may seem surprising that mutations in a gene as highly visible as EGFR and in such a prevalent cancer had not been detected earlier. But the motivation to search aggressively for EGFR mutations was not there until the clinical responses were seen. Perhaps even more surprising was the failure of the pharmaceutical company sponsors of the two most advanced compounds, gefitinib and erlotinib, to embrace this important discovery and refocus future clinical development plans on patients with EGFR mutant lung adenocarcinoma.
But that was 2004, when the prevailing approach to cancer drug development was an empiric one originally developed (with great success) for cytotoxic agents. Typically, small numbers of patients with different cancers were treated in all comer phase I studies (no enrichment for subgroups) with the goal of eliciting a clinical signal in at least one tumor type. A single-agent response rate of 20% to 30% in a disease-specific phase II trial would justify a randomized phase III registration trial, where the typical endpoint for drug approval is time to progression or survival. Cytotoxics were also typically evaluated in combination with existing standard of care treatment (typically approved chemotherapy agents) with the goal of increasing the response rate or enhancing the duration of response. (Note: The use of the past tense here is intentional. As we will see later in this chapter, nearly all cancer drug development today is based on selecting patients with a certain molecular profile.)
The clinical development of gefitinib and erlotinib followed the cytotoxic model. Both drugs had similarly low but convincing single-agent response rates (10% to 15%) in chemotherapy-refractory, advanced lung cancer. Indeed, gefitinib was originally granted accelerated approval by the U.S. Food and Drug Administration (FDA) in 2003 based on the impressive nature of these responses, contingent on the completion of formal phase III studies with survival endpoints.13 The sponsors of both drugs, therefore, conducted phase III registration studies in patients with chemotherapy-refractory, advanced stage lung cancer but without prescreening patients for EGFR mutation status. (In fairness, these trials were initiated prior to the discovery of EGFR mutations in lung cancer but study amendments could have been considered.) Erlotinib was approved in 2004 on the basis of a modest survival advantage over placebo (the BR.21 trial); however, gefitinib failed to demonstrate a survival advantage in essentially the same patient population.14,15 This difference in outcome was surprising because the two drugs have highly similar chemical structures and biologic properties. Perhaps the most important difference was drug dose. Erlotinib was given at the maximum tolerated dose, which produces a high frequency of rash and diarrhea. Both side effects are presumed on target consequences of EGFR inhibition because EGFR is highly expressed in skin and gastrointestinal epithelial cells. In contrast, gefitinib was dosed slightly lower to mitigate these toxicities, with the rationale that responses were clearly documented at lower doses.
In parallel with the single-agent phase III trials in chemotherapy-refractory patients, both gefitinib and erlotinib were studied as an upfront therapy for advanced lung cancer to determine if either would improve the efficacy of standard doublet (carboplatin/paclitaxel or gemcitabine/cisplatin) chemotherapy when all three drugs were given in combination. These trials, termed INTACT-1 and INTACT-2 (gefitinib with either gemcitabine/cisplatin or with carboplatin/paclitaxel) and TRIBUTE (erlotinib with carboplatin/paclitaxel), collectively enrolled over 3,000 patients.16,17,18 Excitement in the oncology community was high based on the clear single-agent activity of both EGFR inhibitors. But, both trials were spectacular failures; neither drug showed any benefit over chemotherapy alone. The fact that EGFR mutations are present in only 10% to 15% of patients (i.e., those likely to benefit) provided a logical explanation. The clinical signal from those whose tumors had EGFR mutations was likely diluted out by all the patients whose tumors had no EGFR alterations, many of whom benefited from chemotherapy.
The convergence of the EGFR mutation discovery with these clinical trial results will be remembered as a remarkable time in the history of targeted cancer therapies, not just for the important role of these agents as lung cancer therapies, but also for missteps in deciding that the EGFR genotype should drive treatment selection. Perhaps the most egregious error came from a retrospective analysis of tumors from patients treated on the BR.21 trial, which concluded that EGFR mutations did not predict for a survival advantage.19 (EGFR gene amplification was associated with survival, but only in a univariate analysis.) This conclusion was concerning because less than 30% of patients on the trial had tissue available for EGFR mutation analysis, raising questions about the adequacy of the sample size. Furthermore, the EGFR mutation assay used by the authors was subsequently criticized because a significant number of the EGFR mutations reported in these patients were in residues not previously found by others, who had sequenced thousands of tumors. Many of these mutations were suspected to be an artifact of working from formalin-fixed biopsies. Fortunately, recent advances in DNA mutation detection, using massively parallel next-generation sequencing technology, have largely eliminated this concern. These new platforms are now being used in the clinical setting.
Clinical investigators in Asia, where a greater fraction of lung cancers (roughly 30%) are positive for EGFR mutations, addressed the question of whether mutations predict for clinical benefit in a prospective trial. In this study known as IPASS, gefitinib was clearly superior to standard doublet chemotherapy as frontline therapy for patients with advanced EGFR mutation-positive lung adenocarcinoma.20 Conversely, EGFR mutation-negative patients fared much worse with gefitinib and benefited from chemotherapy. In addition, EGFR mutation-positive patients had a more favorable overall prognosis regardless of treatment, indicating that EGFR mutation is also a prognostic biomarker. The IPASS trial serves as a compelling example of a properly designed (and executed) biomarker-driven clinical trial. Although the rationale for this clinical development strategy had been demonstrated years earlier with BCR-ABL in leukemia, KIT in GIST, and HER2 in breast cancer, it was difficult to derail the empiric approach that had been used for decades in developing cytotoxic agents.
A Mix of Science and Serendipity: PDGF Receptor-Driven Leukemias and Sarcoma
The discovery of EGFR mutations in lung cancer (motivated by dramatic clinical responses in a subset of patients treated with EGFR kinase inhibitors) is the most visible example of the power of bedside-to-bench science, but it is not the only (or the first) such example from the kinase inhibitor era. Shortly after the approval of imatinib for CML in 2001, two case reports documented dramatic remissions in patients with hypereosinophilic syndrome (HES), a blood disorder characterized by prolonged elevation of eosinophil counts and subsequent organ dysfunction from eosinophil infiltration, when treated with imatinib.21,22 Although HES resembles myeloproliferative diseases such as CML, the molecular pathogenesis of HES was completely unknown at the time. Reasoning that these clinical responses must be explained by inhibition of a driver kinase, a team of laboratory-based physician/scientists quickly searched for mutations in the three kinases known to be inhibited by imatinib (ABL, KIT, and PDGF receptor). ABL and KIT were quickly excluded, but the PDGF receptor α (PDGFR-α) gene was targeted by an interstitial deletion that fused the upstream FIP1L1 gene to PDGFR-α23 FIP1L1-PDGFR-α is a constitutively active tyrosine kinase, analogous to BCR-ABL, and is also inhibited by imatinib. As with EGFR-mutant lung cancer, the molecular pathophysiology of HES was discovered by dissecting the mechanism of response to the drug used to treat it.
The HES/FIP1L1-PDGFR-α story serves as a nice bookend to an earlier discovery that the t(5,12) chromosome translocation, found rarely in patients with chronic myelomonocytic leukemia, creates the TEL-PDGFR-β fusion tyrosine kinase.24 Similar to HES, treatment of patients with t(5,12) translocation-positive leukemias with imatinib has also proven successful.25 A third example comes from dermatofibrosarcoma protuberans, a sarcoma characterized by a t(17,22) translocation that fuses the COL1A gene to the PDGFB ligand (not the receptor). COL1A-PDGFB is oncogenic through autocrine stimulation of the normal PDGF receptor in these tumor cells. Patients with dermatofibrosarcoma protuberans respond to imatinib therapy because it targets the PDGF receptor, just one step downstream from the oncogenic lesion.26
Exploiting the New Paradigm: Searching for Other Kinase-Driven Cancers
The benefits of serendipity notwithstanding, the growing number of examples of successful kinase inhibitor therapy in tumors with a mutation or amplification of the drug target begged for a more rational approach to drug discovery and development. In 2002, the list of human tumors known to have mutations in kinases was quite small. Due to advances in automated gene sequencing, it became possible to ask whether a much larger fraction of human cancers might also have such mutations through a brute force approach. To address this question comprehensively, one would have to sequence all of the kinases in the genome in hundreds of samples of each tumor type. Several early pilot studies demonstrated the potential of this approach by revealing important new targets for drug development. Perhaps the most spectacular was the discovery of mutations in the BRAF kinase in over half of patients with melanoma, as well as in a smaller fraction of colon and thyroid cancers.27 Another was the discovery of mutations in the JAK2 kinase in nearly all patients with polycythemia vera, as well as a significant fraction of patients with myelofibrosis and essential thrombocytosis.28,29,30 A third example was the identification of PIK3CA mutations in a variety of tumors, with the greatest frequencies in breast, endometrial, and colorectal cancers.31 PIK3CA encodes a lipid kinase that generates the second messenger phosphatidyl inositol 3-phosphate (PIP3). PIP3 activates growth and survival signaling through the AKT family of kinases as well as other downstream effectors. Coupled with the well-established role of the phosphatase and tensin homolog (PTEN) lipid phosphatase in dephosphorylating PIP3, the discovery of PIK3CA mutations focused tremendous attention on developing inhibitors at multiple levels of this pathway, as discussed further in the follow paragraphs.
Each of these important discoveries—BRAF, JAK2, and PIK3CA—came from relatively small efforts (less than 100 tumors) and generally focused on resequencing only those exons that coded for regions of kinases where mutations had been found in other kinases (typically, the juxtamembrane and kinase domains). These restricted searches were largely driven by the high cost of DNA sequencing using the Sanger method. In 2006, a comprehensive effort to sequence all of the exons in all kinases in 100 tumors could easily exceed several million dollars. Financial support for such projects could not be obtained easily through traditional funding agencies because the risk/reward was considered too high. Furthermore, substantial infrastructure for sample acquisition, microdissection of the tumors from normal tissue, nucleic acid preparation, high throughput automated sequencing, and computational analysis of the resulting data was essential. Few institutions were equipped to address these challenges. In response, the National Cancer Institute in the United States (in partnership with the National Human Genome Research Institute) and an international group known as the International Cancer Genome Consortium (ICGC) launched large-scale efforts to sequence the complete genomes of thousands of cancers. In parallel, next-generation sequencing technologies resulted in massive reductions in cost, allowing a more comprehensive analysis of much larger numbers of tumors. At the time of this writing, the US effort (called The Cancer Genome Atlas [TCGA]) had reported data on 29 different tumor types (https://tcga-data.nci.nih.gov/tcga/). The international consortium has committed to sequencing 25,000 tumors representing 50 different cancer subtypes.32 Both groups have enforced immediate release of all sequence information to the research community free of charge so that the entire scientific community can learn from the data. This policy enabled pan cancer mutational analyses that give an overall view of the genomic landscape of cancer, serving as a blueprint for the community of cancer researchers and drug developers.33,34
Rounding Out the Treatment of Myeloproliferative Disorders: JAK2 and Myelofibrosis
Taken together with the BCR-ABL translocation in CML and FIP1L1-PDGFR-α in HES, the discovery of JAK2 mutations in polycythemia, essential thrombocytosis, and myelofibrosis provided a unifying understanding of myeloproliferative disorders as diseases of abnormal kinase activation. The JAK family kinases are the primary effectors of signaling through inflammatory cytokine receptors and, therefore, had been considered compelling targets for anti-inflammatory drugs. But the JAK2 mutation discovery immediately shifted these efforts toward developing JAK2 inhibitors for myeloproliferative disorders. Because most patients have a common JAK2 V617F mutation, these efforts could rapidly focus on screening for activity against a single genotype. Progress has been rapid. Myelofibrosis was selected as the initial indication (instead of essential thrombocytosis or polycythemia vera) because the time to registration is expected to be the shortest. Currently, ruxolitinib is approved for myelofibrosis based on shrinkage in spleen size as the primary endpoint. Clinical trials in essential thrombocytosis and polycythemia vera (versus hydroxyurea) are ongoing. Other JAK2 inhibitors are also in clinical development.
BRAF Mutant Melanoma: Several Missteps Before Finding the Right Inhibitor
As with JAK2 mutations in myeloproliferative disorders, the discovery of BRAF mutations in patients with melanoma launched widespread efforts to find potent BRAF inhibitors. One early candidate was the drug sorafenib, which had been optimized during drug discovery to inhibit RAF kinases. (Sorafenib also inhibits vascular endothelial growth factor (VEGF) receptors, which led to its approval in kidney cancer, as discussed later in this chapter.) Despite the compelling molecular rationale for targeting BRAF, clinical results of sorafenib in melanoma were extremely disappointing and reduced enthusiasm for pursuing BRAF as a drug target.35 In hindsight, this concern was completely misguided. Sorafenib dosing is limited by toxicities that preclude achieving serum levels in patients that potently inhibit RAF, but are sufficient to inhibit VEGF receptors. In addition, patients were enrolled without screening for BRAF mutations in their tumors. Although the frequency of BRAF mutations in melanoma is high, the inclusion of patients without the BRAF mutation diluted the chance of seeing any clinical signal. In short, the clinical evaluation of sorafenib in melanoma was poorly designed to test the hypothesis that BRAF is a therapeutic target. The danger is that negative data from such clinical experiments can slow subsequent progress. It is critical to know the pharmacodynamic properties of the drug and the molecular phenotype of the patients being studied when interpreting the results of a negative study.
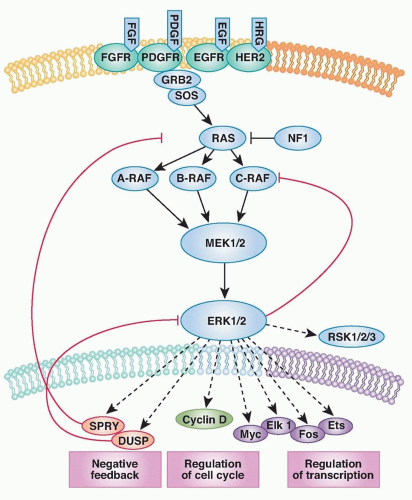
The fact that RAF kinases are intermediate components of the well-characterized RAS/mitogen-activated protein (MAP) kinase pathway (transducing signals from RAS to RAF to MEK to ERK) raised the possibility that tumors with BRAF mutations might respond to inhibitors of one of these downstream kinases (Fig. 22.1). Preclinical studies revealed that tumor cell lines with BRAF mutation were exquisitely sensitive to inhibitors of the downstream kinase MEK.36 (Sorafenib, in contrast, does not show this profile of activity.37 Thus, proper preclinical screening would have revealed the shortcomings of sorafenib as a BRAF inhibitor.) Curiously, cell lines with a mutation or amplification of EGFR or HER2, which function upstream in the pathway, were insensitive to MEK inhibition. Even tumor lines with RAS mutations were variably sensitive. In short, the preclinical data made a strong case that MEK inhibitors should be effective in BRAF mutant melanoma, but not in other subtypes. The reason that HER2, EGFR, and RAS mutant tumors were not sensitive to MEK inhibitors is explained, at least in part, by the existence of negative feedback loops that modulate the flux of signal transduction through MEK.38
Figure 22.1 The RAS-RAF-MEK-ERK signaling pathway. The classical mitogen-activated protein kinase (MAPK) pathway is activated in human tumors by several mechanisms, including the binding of ligand to receptor tyrosine kinases (RTK), the mutational activation of an RTK, by loss of the tumor suppressor NF1, or by mutations in RAS, BRAF, and MEK1. Phosphorylation and, thus, activation of ERK regulates the transcription of target genes that promote cell cycle progression and tumor survival. The ERK pathway contains a classical feedback loop in which the expression of feedback elements such as SPRY and DUSP family proteins are regulated by the level of ERK activity. Loss of expression of SPRY and DUSP family members due to promoter methylation or deletion is thus permissive for persistently elevated pathway output. In the case of tumors with mutant BRAF, pathway output is enhanced by impaired upstream feedback regulation. FGF, fibroblast growth factor; HRG, heregulin; NF1, neurofibromatosis 1. (From Bernt KM, Zhu N, Sinha AU, et al. MLL-rearranged leukemia is dependent on aberrant H3K79 methylation by DOT1L. Cancer Cell 2011;20(1):66-78, with permission.)
In parallel with the generation of these preclinical findings, clinical trials of several MEK inhibitors were initiated. Patients with various cancers were enrolled in the early studies, but there was a strong bias to include melanoma patients. Significant efforts were made to demonstrate MEK inhibition in tumor cells by measuring the phosphorylation status of the direct downstream substrate ERK using an immunohistochemical analysis of biopsies from patients with metastatic disease. Phase I studies of the two earliest compounds in clinical development (PD325901 and AZD6244) documented reduced phospho-ERK staining at multiple dose levels in several patients for whom baseline and treatment biopsies were obtained.39,40 (In the following, we will learn that these pharmacodynamic studies, while well intentioned, were not quantitative enough to document the magnitude of MEK inhibition in these patients.) Furthermore, clinical responses were observed in a few patients with BRAF mutant melanoma. Armed with this confidence, a randomized phase II clinical trial of AZD6244 was conducted in advanced melanoma, with the chemotherapeutic agent temozolomide (which is approved for glioblastoma) as the comparator arm. (The clinical development of PD325901 was discontinued because of safety concerns about ocular and neurologic toxicity.) Disappointingly, patients receiving AZD6244 had no benefit in progression-free survival when compared to temozolomide-treated patients, raising further concerns about the viability of BRAF as a drug target.41 A closer examination of the data revealed that clinical responses were, indeed, seen in patients receiving AZD6244. The fact that BRAF mutation status was not required for study entry likely diminished the clinical signal in the AZD6244 arm, a lesson learned from the EGFR inhibitor trials in lung cancer. Indeed, a different MEK inhibitor, trametinib, received FDA approval in 2013 based on activity in melanoma patients with the BRAF mutation.42
Only gold members can continue reading. Log In or Register to continue