Fc Receptors and Their Role in Immune Regulation and Inflammation
Jeffrey V. Ravetch
Falk Nimmerjahn
HISTORICAL BACKGROUND
Cellular receptors for immunoglobulins (Igs) were anticipated by the description of cytophilic antibodies of the IgG class, identified by Boyden and Sorkin in 1960.1 These antibodies conferred upon normal cells, like macrophages, the capacity to specifically absorb antigen. Using sheep red blood cells (RBCs) as the antigen resulted in rosette formation between the cytophilic antisheep RBC antibodies and macrophages and provided a convenient means of visualization of the binding of cytophilic antibodies with normal cells. Subsequent studies by Berken and Benacerraf2 suggested that the crystallized fragment (Fc) of the cytophilic antibody interacted with a cell surface receptor on macrophages. Similar studies on B-lymphocytes extended the generality of these receptors and led to the term Fc receptor (FcR) to denote the surface molecules on lymphoid and myeloid cells that are capable of interacting with the Fc of immunoglobulin molecules.3 Studies on IgE, IgM, and IgA demonstrated the existence of distinct receptors for those isotypes as well on various immune cell types. Detailed biochemical characterization of Fc receptors was inaugurated by the studies of Kulczycki et al.4 on the high-affinity IgE FcR of mast cells, revealing a hetero-oligomeric &agr;&bgr;&ggr;2 subunit structure. A distinction between FcRs for the IgE and IgG isotypes emerged with the observation of the very high (1010 M-1) binding affinity of IgE for its receptor in comparison with the low binding (106 M-1) of IgG1 to its receptor. This distinction led to the realization that the functional IgG1 ligand was exclusively in the form of an immune complex (IC), whereas IgE binding occurred through monomer interaction with its receptor. This difference in binding affinity had significant functional implications for the structures of these receptors and mechanisms by which each isotype activated its target cell. Determination of the structure of these receptors was facilitated by their molecular cloning, beginning with the IgG FcRs5,6 and followed by the IgE FcR.7 Two distinct types of IgG receptors, differing in their transmembrane and cytoplasmic sequences, were identified that offered a molecular explanation for the apparent contradictory activation and inhibitory activities attributed to IgG FcRs. The primary structure of the subunits of the high-affinity IgE FcR revealed homology in the ligand binding &agr; subunit to its IgG counterparts. However, the extent of similarity between these receptors became apparent with the observation that the &ggr; chain subunit was common to both IgG and IgE FcRs, providing both assembly and signaling functions to these activation receptors.8,9 This common structure suggested a functional link between IC diseases and allergic reactions, a prediction that was confirmed through mouse knockout studies of IgG FcRs.10,11 The FcRs, through their dependence on the immunoreceptor tyrosine-based activation motif (ITAM) pathway of cellular activation, belonged to the family of immunoreceptors that included the antigen receptors on B and T cells. Three-dimensional crystal structures have been solved for the low-affinity IgG FcRs12,13 and the high-affinity IgE FcR,14 alone and in complex with their Ig ligands,15,16 further establishing the close structural link between these Ig receptors.
The functional roles of IgG FcRs were suggested by the distribution of these receptors on both lymphoid and myeloid cells.17 On myeloid cells, they were presumed to mediate effector cell activation, resulting in phagocytosis, antibody-dependent cellular cytotoxicity (ADCC), and release of inflammatory mediators. However, the well-known ability of the classical pathway of complement to generate activated fragments in response to ICs capable of inducing inflammatory responses by myeloid cells complicated the interpretation of the physiological role of IgG FcRs. Thus, the contribution of IgG FcRs to the mechanism of IC-mediated inflammation, as distinct from the role of complement, remained uncertain. Insight into this distinction was gained through the generation of mouse strains specifically deficient or blocked in either FcRs10,18,19,20 or components of the classical complement pathway.21 Studies on IC-mediated inflammatory responses in these animals, such as the Arthus reaction, led to the realization that IgG FcRs and not the classical pathway of complement activation were the functional mediators of inflammatory responses triggered by ICs.20,22,23 The situation for IgE was less confounding, and the identification and characterization of a high-affinity receptor for this isotype on mast cells offered a plausible explanation for many of the inflammatory features of allergic reactions,24 validated later by mouse knockouts of this receptor. IgG ICs had also been observed to mediate suppression of B-cell responses; thus, the presence of an IgG FcR activity on B cells provided a possible, but uncharacterized, mechanism for this inhibitory activity. Molecular characterization of this inhibitory activity for the B-cell FcR, FcRIIB, resulted in the first detailed description of an inhibitory motif, now termed the immunoreceptor tyrosine-based inhibitory motif (ITIM),25,26 and the signaling pathway by which it abrogates ITAM-triggered activation. The ITIM mechanism is now recognized as ubiquitous and has resulted in the recognition of a large family of inhibitory receptors on immune cells that function to maintain proper
thresholds for activation and abrogate activation responses to terminate an immune reaction.27
thresholds for activation and abrogate activation responses to terminate an immune reaction.27
FcRs are now recognized as central mediators of antibody-triggered responses, coupling the innate and adaptive immune responses in effector cell activation.28 In addition to these specialized roles, the IgG FcRs have served as an example of the emerging class of balanced immunoreceptors, in which activation and inhibition are tightly coupled in response to ligand binding. Perturbations in either arm of the response have been shown to lead to pathological consequences and have been taken as a paradigm of how these systems are likely to work for those paired immunoreceptors with unknown ligand-binding functions. The newly described roles for FcRs in maintaining peripheral tolerance, shaping the antibody repertoire, regulating antigen-presenting cell (APC) maturation, and promoting mast cell survival indicate the diversity of functions that these receptors possess and their central role in modulating both afferent and efferent responses in the immune response.
This chapter focuses primarily on the IgG and IgE FcRs, for which substantial data on their structure, function, regulation, and role in a variety of physiological and pathological conditions are now available. The similarity in structure and signaling between those receptors and other members of this family, such as the IgA FcR and the recently discovered Fc-receptor for mouse and human IgM (Fc&mgr;R), is also discussed. These FcRs will be referred to as “canonical Fc receptors,” reflecting their shared structural and functional properties. In contrast, lectins such as SIGN-R1 and DC-SIGN, which bind IgG Fc when sialylated, represent a novel class of FcRs with distinct structures and functions and will be discussed in detail in the following. Other Ig receptors with specialized functions in the transport of Igs, such as the FcRn29 and the poly-Ig FcR,30 will not be discussed here.
STRUCTURE AND EXPRESSION
Molecular Genetics
Two general classes of canonical FcRs are now recognized: the activation receptors, characterized by the presence of a cytoplasmic ITAM sequence associated with the receptor, and the inhibitory receptor, characterized by the presence of an ITIM sequence. These two classes of receptors function in concert and are usually found coexpressed on the cell surface. Thus coengagement of both signaling pathways is the rule, setting thresholds for and ultimately determining the physiological outcome of effector cell responses. Among the factors that determine this threshold level are the actual affinities of the individual activating and inhibitory receptors for a specific IgG ligand and the expression level of the receptor pairs on immune effector cells. Importantly, the affinity of different antibody isotypes and subclasses for their respective activating and inhibitory FcRs varies significantly, thus explaining the differential activity of antibody isotypes in vivo.20,31,32 In addition, alleles of specific FcRs have been described that alter their affinity for individual subclasses thus accounting for the variable responses seen in a population to an antibody (see subsequent discussion). Noncanonical FcRs such as the IgM receptors Fc&mgr;R, Fca/&mgr;R, the poly-Ig receptor, and the sialylated IgG Fc receptors SIGN-R1 and DC-SIGN, contain neither ITAM nor ITIM motifs.
Subunit Composition
Canonical FcRs are typically type I integral membrane glycoproteins consisting of, at the least, a ligand recognition &agr; subunit that confers isotype specificity for the receptor. &agr; subunits for IgG, IgE, IgM, and IgA have been described.17,33,34,35 These subunits typically consist of two extracellular domains of the IgV type superfamily, a single transmembrane domain and a relatively short intracytoplasmic domain. In activation FcRs, a signaling subunit of the &ggr; family is often found, resulting in an &agr;&ggr;2 complex. The inhibitory Fc&ggr;RIIB molecule, in contrast, is expressed as a single-chain receptor. The &agr; subunits have apparent molecular weights of between 40 and 75 kDa, and share significant amino acid sequence homology in their extracellular domains. Alternatively, spliced forms of Fc&ggr;RIIB modify the intracytoplasmic domain of this molecule. For example, the B2 form lacks sequences that inhibit internalization and thus demonstrates enhanced internalization of ICs, in comparison with Fc&ggr;RIIB1. However, all the splice variants contain the ITIM motif, a necessary and sufficient domain for mediating inhibitory signaling. The conservation of this sequence in mice and humans, its presence in all splice variants, and the hyperresponsive phenotypes generated in mice deficient in this receptor all support inhibition as the central function of Fc&ggr;RIIB. The specific structures of the &agr; subunits of the canonical FcRs are shown in Figure 24.1. The notable exceptions to the general structure just outlined are seen for the high-affinity Fc&ggr;RI&agr; subunit, which has three extracellular domains; the activation Fc&ggr;RIIA&agr; subunit, which does not require additional subunits for assembly or signaling; and the glycosylphosphatidyl-inositol (GPI)-linked Fc&ggr;RIIIB, which attaches to the cell surface through a GPI linkage, rather than through a transmembrane domain.
The &ggr; subunit is found associated with activation IgG, IgE, and IgA FcRs, as well as with non-FcR molecules, such as paired Ig-like receptor A (PIR-A) and natural killer (NK) cell cytotoxicity receptors, but not with IgM receptors, nor the lectins SIGN-R1 and DC-SIGN.36,37,38 It is required for assembly of the &agr; subunits of these receptors by protecting these subunits from degradation in the endoplasmic reticulum. The &ggr;chain is found as a disulfide-linked homodimer, with a short extracellular domain containing the cysteine involved in dimerization, a transmembrane domain, and an intracytoplasmic domain containing the ITAM. An aspartic acid residue found in the transmembrane domain of the &ggr;chain is often associated with a basic amino acid residue in the transmembrane domain of the &agr; subunit. The &ggr;subunit belongs to a gene family that includes the T-cell receptor-associated &zgr; chain and the NK receptor DAP-10- and DAP-12-associated molecules.39 Fc&ggr;RIIIA can associate with the &zgr; chain, resulting in the &agr;&zgr;2 complex found in human NK cells.
A third subunit is found associated with the activation FcRs Fc&egr;RI and Fc&ggr;RIII, the &bgr; subunit. This 33-kDa subunit has four transmembrane-spanning domains and amino and carboxy intracytoplasmic domains, belonging to the cluster
of differentiation (CD)20 family of tetraspan molecules.24 An ITAM sequence is found in the intracytoplasmic carboxy domain. In mast cells and basophils, the &bgr; chain assembles into an &agr;&bgr;&ggr;2 complex with the &agr; chain belonging to either Fc&ggr;RIII or Fc&egr;RI. Its presence is required for assembly of Fc&egr;RI in rodents. In humans, however, &agr;&ggr;2 complexes of Fc&egr;RI are found in monocytes, Langerhans cells, and dendritic cells (DCs), in addition to the &agr;&bgr;&ggr;2 complexes found in mast cells and basophils. The ITAM motif found in the &bgr; subunit is not an autonomous activation sequence but functions as a signaling amplifier of the ITAM found in the &ggr;subunits.40
of differentiation (CD)20 family of tetraspan molecules.24 An ITAM sequence is found in the intracytoplasmic carboxy domain. In mast cells and basophils, the &bgr; chain assembles into an &agr;&bgr;&ggr;2 complex with the &agr; chain belonging to either Fc&ggr;RIII or Fc&egr;RI. Its presence is required for assembly of Fc&egr;RI in rodents. In humans, however, &agr;&ggr;2 complexes of Fc&egr;RI are found in monocytes, Langerhans cells, and dendritic cells (DCs), in addition to the &agr;&bgr;&ggr;2 complexes found in mast cells and basophils. The ITAM motif found in the &bgr; subunit is not an autonomous activation sequence but functions as a signaling amplifier of the ITAM found in the &ggr;subunits.40
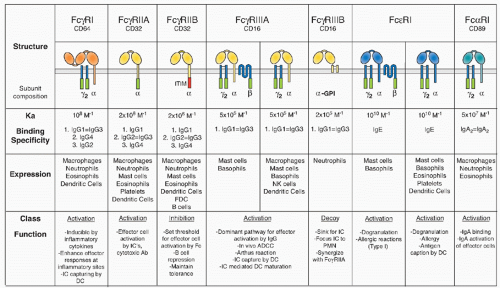 FIG. 24.1. Summary of Fc Receptor Structures, Expression Patterns, and in Vivo Functions. The immunoreceptor tyrosine-based activation motif signaling motif is indicated by the green rectangle; the immunoreceptor tyrosine-based inhibitor motif is indicated as a red rectangle. Alleles of Fc&ggr;RIIA and Fc&ggr;RIIIB and their binding properties are discussed in the text. |
Gene Organization, Linkage, and Polymorphisms
All &agr; subunits of canonical FcRs share a common gene organization, which indicates that the evolution of this family of receptors resulted from gene duplication from a common ancestor.41 Sequence divergence then resulted in the acquisition of distinctive specificities for these related sequences. Most of the genes belonging to the expanded FcR family, including the recently identified Fc&mgr;-receptor, the Fc&agr;/&mgr;R, the poly-Ig receptor, the group of FcR-homologous or -like (FCRL) proteins, the &agr; chains of Fc&ggr;RI, Fc&ggr;RII, Fc&ggr;RIII, and Fc&egr;RI including the common FcR-&ggr; chain (Fig. 24.2), are found on the long arm of chromosome 1.34,35,42,43,44 This region is syntenic with a comparable region on mouse chromosome 1; however, Fc&ggr;RI&agr; and several FCRLs are found on mouse chromosome 3. In humans, the &agr; subunit of the IgA receptor is found on chromosome 19 as is the lectin DC-SIGN (CD209) and the &bgr; subunit is on chromosome 11. The Fc&ggr;RII-Fc&ggr;RIII locus on chromosome 1 is further linked to a variety of lupus susceptibility genes found in that region, including the Sle1 cluster.45 A locus linked to atopy has been identified at 11q12-13 and further delineated polymorphisms of the &bgr; chain (I181V and V183L) that are associated with a heightened risk of atopy. However, a direct functional association of these polymorphisms with the known biological activities of the &bgr; chain has not been found.46
Polymorphisms in the &agr; chains of the Fc&ggr;Rs have been described, most notably in Fc&ggr;RIIA and Fc&ggr;RIIIA; these polymorphisms result in differences in binding affinity to specific IgG subclasses.47 For example, a histidine at position 131 in Fc&ggr;RIIA results in higher affinity binding to IgG2 and IgG3 than does an arginine at that position. Similarly, Fc&ggr;RIIIA with valine at position 158 of the &agr; chain has a higher binding affinity for IgG1 and 3 than does the polymorphic form with phenylalanine at that position. These polymorphisms translate into a more robust ADCC response for the Fc&ggr;RIIIA val/val and the Fc&ggr;RIIA his/his haplotype in vitro and has been positively correlated with a better clinical response to antitumor antibodies inclunding the CD20-specific antibody rituximab, the Her2/neu-specific antibody trastuzumab, and the epidermal growth factor receptor-specific antibody cetuximab in human lymphoma, metastatic breast cancer, and colorectal cancer patient cohorts, respectively.48,49,50,51,52,53 This data is consistent with previous results obtained with rituximab
and trastuzumab in mouse models, demonstrating an important role of cellular Fc&ggr;Rs for the activity of these therapeutic antibodies in vivo.54,55 Four amino acids are polymorphic for Fc&ggr;RIIIB at positions 18, 47, 64, and 88, which contribute to the neutrophil antigen polymorphisms for this receptor. Several studies have attempted to link specific FcR polymorphisms or copy number variations to autoimmune diseases, specifically to systemic lupus erythematosus (SLE).56 Recent studies have reported associations in susceptibility to SLE in both murine and human populations with levels of Fc&ggr;RIIB
expression or alleles of Fc&ggr;RIIB.57 Reduced expression of Fc&ggr;RIIB on activated B cells, such as memory cells, has been seen in patients with SLE and chronic inflammatory demyelinating polyneuropathy (CIDP), and is associated with a promoter polymorphism.58,59 In mouse strains that develop a spontaneous, lupus-like disease, restoring the level of Fc&ggr;RIIB expression on their B cells to a wild-type level reverses disease.60,61 A higher incidence of an allele of Fc&ggr;RIIB has been reported in several populations with autoimmune disease. This allele, found in the transmembrane domain of the receptor, is suggested to result in a hypomorphic phenotype, similar to the reduced expression observed.57 Of note, despite increasing the risk for development of SLE, this Fc&ggr;RIIB allele seems to decrease the likelihood of malaria infections and the severity of disease and can be found at increased frequencies in areas of the world where malaria is endemic.62,63
and trastuzumab in mouse models, demonstrating an important role of cellular Fc&ggr;Rs for the activity of these therapeutic antibodies in vivo.54,55 Four amino acids are polymorphic for Fc&ggr;RIIIB at positions 18, 47, 64, and 88, which contribute to the neutrophil antigen polymorphisms for this receptor. Several studies have attempted to link specific FcR polymorphisms or copy number variations to autoimmune diseases, specifically to systemic lupus erythematosus (SLE).56 Recent studies have reported associations in susceptibility to SLE in both murine and human populations with levels of Fc&ggr;RIIB
expression or alleles of Fc&ggr;RIIB.57 Reduced expression of Fc&ggr;RIIB on activated B cells, such as memory cells, has been seen in patients with SLE and chronic inflammatory demyelinating polyneuropathy (CIDP), and is associated with a promoter polymorphism.58,59 In mouse strains that develop a spontaneous, lupus-like disease, restoring the level of Fc&ggr;RIIB expression on their B cells to a wild-type level reverses disease.60,61 A higher incidence of an allele of Fc&ggr;RIIB has been reported in several populations with autoimmune disease. This allele, found in the transmembrane domain of the receptor, is suggested to result in a hypomorphic phenotype, similar to the reduced expression observed.57 Of note, despite increasing the risk for development of SLE, this Fc&ggr;RIIB allele seems to decrease the likelihood of malaria infections and the severity of disease and can be found at increased frequencies in areas of the world where malaria is endemic.62,63
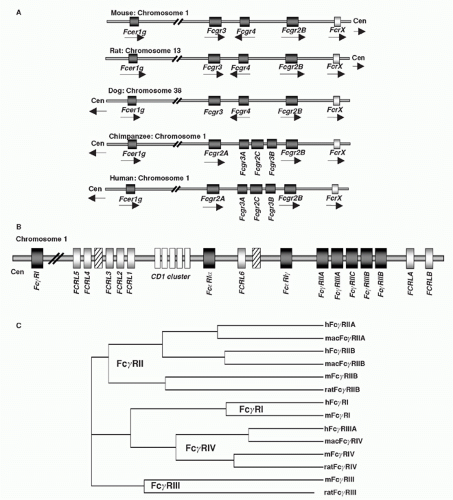 FIG. 24.2. Chromosomal Organization of Fc Receptor (FcR) and FcR-Like Genes. A: Localization of FcR genes in different species. B: The human FcR locus on chromosome 1. Classical FcR genes are shown in dark grey, FCRL genes as light grey, and the CD1 gene cluster on chromosome 1 as white boxes. Pseudogenes are indicated as hatched boxes. (adapted from Davis et al.44). C: The cladogramm shows the alignment of selective classical FcRs of humans (h), macaques (mac), mice (m), and rats. |
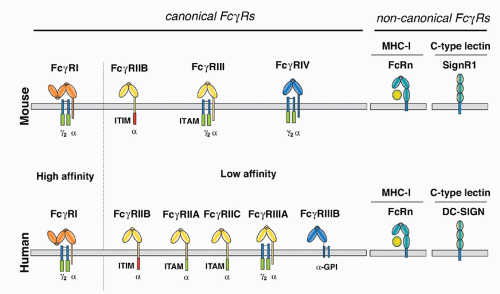 FIG. 24.3. Comparison of the Human and Mouse Canonical and Noncanonical Fc&ggr;-Receptor Protein Family. In both species, canonical Fc&ggr;Rs can be distinguished by their affinity for the antibody Fc-portion (high or low affinity) and by the signaling pathways they trigger (activating versus inhibitory). The noncanonical neonatal FcR belongs to the family of major histocompatibility class I proteins and regulates immunoglobulin (Ig)G half life. Mouse SIGNR1 or human DC-SIGN are C-type lectin proteins that can only bind to IgG glycoforms rich in terminal sialic acid residues and are involved in the anti-inflammatory activity of IgG. |
Species Comparisons
Detailed comparisons between the canonical FcRs in mice and humans have revealed several notable differences in both structure and expression of these molecules (Figs. 24.1, 24.2, 24.3). Whereas IgG, IgM, and IgE FcRs are conserved in these species, IgA FcRs are not. To date, a murine homolog for the IgA FcR has not been identified. In general, murine and human IgG FcRs display comparable degrees of heterogeneity and complexity.41,64 Specific differences, however, have been noted. For example, Fc&ggr;RI is encoded by a single gene in the mouse, in comparison with three genes in the human.43 Two genes for activation FcRs, Fc&ggr;RIIA and C, are found in the human and not rodents, which is notable because of their unusual single-chain activation structure.65 Both mice and human encode a gene referred to as Fc&ggr;RIIIA, although recent studies have identified a novel mouse FcR with higher homology to the human Fc&ggr;RIIIA called Fc&ggr;RIV.31,64,66 As mentioned previously, Fc&ggr;RIIIB is unique among FcRs in being expressed as a GPI-anchored protein (see Fig. 24.3). Its expression is limited to human neutrophils, in comparison with Fc&ggr;RIIIA, which is expressed widely on cells of the myeloid lineage, such as macrophages, NK cells, mast cells, and DCs. Finally, both mice and humans have only a single gene encoding the inhibitory Fc&ggr;RIIB molecule. Among the noncanonical FcRs, the lectin DC-SIGN is the functional homologue of the murine SIGN-R1 molecule with respect to its ability to bind sialylated IgG Fc and mediate an antiinflammatory response.67,68 While the overall structure of these proteins are similar (see Fig. 24.3), their patterns of expression are quite distinct. SIGN-R1 expression is restricted to marginal zone macrophages of the spleen and lymph node, whereas DC-SIGN expression is seen on myeloid DCs and macrophages in a variety of tissues, including spleen, bone marrow, and lymph node.69
The genes for the IgE FcR are conserved in mice and humans. The difference that is observed relates to the requirement for the &bgr; chain to achieve surface expression
in mice, precluding the expression of the &agr;&ggr;2 complex.24 In humans, this form of the receptor is widely expressed on monocytes, Langerhans cells, and DCs, and is likely to be found on mast cells and basophils as well. This difference in Fc&egr;RI subunit composition is likely to result in functional differences as well. Although these specific interspecies differences are important, the fundamental organization of the canonical FcR system, with activation and inhibitory signaling through a shared ligand specificity coupled to opposing signaling pathways, is well conserved, as is the role of the noncanonical FcRs in mediating the immunomodulatory functions of sialylated IgG. Thus, conclusions regarding the function of this expanded FcR system in immunity by the analysis of murine models are relevant to an understanding of the role of these receptors to human immunity as well.
in mice, precluding the expression of the &agr;&ggr;2 complex.24 In humans, this form of the receptor is widely expressed on monocytes, Langerhans cells, and DCs, and is likely to be found on mast cells and basophils as well. This difference in Fc&egr;RI subunit composition is likely to result in functional differences as well. Although these specific interspecies differences are important, the fundamental organization of the canonical FcR system, with activation and inhibitory signaling through a shared ligand specificity coupled to opposing signaling pathways, is well conserved, as is the role of the noncanonical FcRs in mediating the immunomodulatory functions of sialylated IgG. Thus, conclusions regarding the function of this expanded FcR system in immunity by the analysis of murine models are relevant to an understanding of the role of these receptors to human immunity as well.
Expression
Canonical FcRs are expressed widely on cells of the myeloid lineage, including monocytes, macrophages, DCs, mast cells, basophils, neutrophils, eosinophils, and NK cells.17,33 In addition, B cells and follicular DCs (FDCs) express the inhibitory Fc&ggr;RIIB receptor, whereas T cells are generally negative for Fc&ggr;R expression, but may express the Fc-receptor for IgM. Interestingly, the majority of FCRL proteins are expressed during varying stages of B-cell development. Despite their homology to the canonical FcRs, however, the FCRL proteins seem not to bind to Igs, rendering Fc&ggr;RIIB the only IgG binding FcR on B cells. The specific expression pattern for each FcR varies, and these patterns are summarized in Figure 24.1. Because canonical FcRs represent a balanced system of activation and inhibition, the general rule of coexpression of FcRs of these classes is maintained. B cells use the B-cell antigen receptor as the activation coreceptor for Fc&ggr;RIIB, whereas NK cells appear to utilize NK inhibitory receptors to modulate Fc&ggr;RIIIA activation. The decoy Fc&ggr;R, Fc&ggr;RIIIB, is expressed exclusively on human neutrophils, on which it functions to concentrate and focus ICs without directly triggering cell activation, perhaps also playing role in neutrophil recruitment.70 The Fc&ggr;RIIA-Fc&ggr;RIIB pair functions on neutrophils to modulate IC activation. Fc&egr;RI can be modulated by Fc&ggr;RIIB, as demonstrated both in vitro and in vivo; mice deficient in Fc&ggr;RIIB display enhanced IgE-triggered anaphylaxis71 by virtue of the ability of IgE to bind with high affinity to Fc&egr;RI and with low affinity for Fc&ggr;RIIB. Other mast cell inhibitory receptors, such as glycoprotein 49B1, modulate mast cell sensitivity to IgE: mice deficient in this molecule display enhanced anaphylactic responses to IgE stimulation.72 Expression of the common &ggr; chain is broad: it has been found on all myeloid and lymphoid cells examined to date. In contrast, the &bgr; chain appears to be quite restricted in its expression: it has been found only on mast cells and basophils.
Regulation of canonical FcR expression can occur at several levels. In general, cytokines involved in activation of inflammatory responses induce expression of activation Fc&ggr;Rs, whereas inhibitory cytokines downregulate these activation receptors. Transcriptional regulation of &agr; chain levels has been documented for a variety of cytokines, including interferon-&ggr;, interleukin (IL)-4, and transforming growth factor &bgr;.73,74 In addition, complement component C5a binding to its receptor, C5aR, results in the induction of expression of activation Fc&ggr;Rs.75 Induction of Fc&ggr;RI, Fc&ggr;RIIA, and Fc&ggr;RIIIA &agr; and &ggr;chains in myeloid cells occurs upon interferon-&ggr;treatment: IL-4 generally inhibits expression of these activation receptors but induces expression of the inhibitory Fc&ggr;RIIB. Administration of intravenous gamma globulin, a widely used treatment for inflammatory diseases, has been shown to induce expression of the inhibitory Fc&ggr;RIIB on effector macrophages and B cells in mice and humans.59,76,77,78 This induction is not direct but mediated through other, noncanonical FcR pathways including molecules such as SIGNR1 and its human orthologe DC-SIGN (see Fig. 24.7). Engagement of these receptors by sialylated IgG results in an intrinsic Th2 pathway, ultimately leading to the expression of IL-4, which induces FcRIIB expression on inflammatory macrophages.67,68,79,80 The situation in B cells is likely to be more complex, whereby regulation of Fc&ggr;RIIB is critical for the maintenance of peripheral tolerance. Germinal center B cells downregulate Fc&ggr;RIIB, perhaps in response to IL-4 production by T cells. Regulation of FcR expression has also been documented to occur upon binding of ligand. IgE regulates the expression of Fc&egr;RI by stabilizing the intracellular pool of receptor upon receptor engagement.81 Thus, high IgE levels result in the induction of surface expression of Fc&egr;RI. However, this same mechanism of regulation is not seen for Fc&ggr;Rs: mice deficient in IgG have Fc&ggr;R levels comparable with those of wild-type animals. Competition for limiting subunits also contributes to regulation of receptor expression. In mast cells, it appears that the level of &ggr; chain is limiting. Competition between &agr; chains for the limiting concentration of &ggr; chain has been documented in mouse knockouts, whereby levels of one receptor increase if the &agr; chain of the other receptor is reduced.82 This type of reciprocal regulation has recently also been observed for Fc&ggr;Rs in neutrophils and is likely to be significant in the cross-regulation of FcRs by different isotypes of Ig.83
Three-Dimensional Structure
The crystal structures of Fc&ggr;RIIA, Fc&ggr;RIIB, Fc&ggr;RIIIA, and Fc&egr;RI have been solved, as have the cocrystals of Fc&ggr;RIIIA-IgG1 Fc and Fc&egr;RI-IgE Fc84 (Fig. 24.4). These studies demonstrate that the receptors have a common structure in which the two extracellular Ig domains fold in a strongly bent overall structure, arranged into a heart-shaped domain structure. A 1:1 stoichiometry between the receptor and ligand is observed, with the receptor inserted into the cleft formed by the two chains of the Fc fragment (C&ggr;2 or C&egr;3). The binding region of the FcR to Fc fragments consists mainly of rather flexible loops that rearrange upon complex formation. Only domain 2 and the linker region connecting domains 1 and 2 interact in the complex with different regions of both chains of the Fc. Conserved tryptophans located on the FcRs interact with proline to form a “proline sandwich.” A solvent-exposed hydrophobic residue at position 155 is conserved among all FcRs and represents a binding site for the important IgG1 residue Leu 235 (not
found in IgE). Specificity is generated among the receptor-ligand pairs in a variable region connecting the two extracellular domains that is in contact with the lower hinge region of the Fc fragment (residues 234 to 238), a region not conserved among the IgGs and IgE. The binding region of FcRs to their Ig ligands does not overlap with other Fc binding molecules such as protein A, protein G, and FcRn.
found in IgE). Specificity is generated among the receptor-ligand pairs in a variable region connecting the two extracellular domains that is in contact with the lower hinge region of the Fc fragment (residues 234 to 238), a region not conserved among the IgGs and IgE. The binding region of FcRs to their Ig ligands does not overlap with other Fc binding molecules such as protein A, protein G, and FcRn.
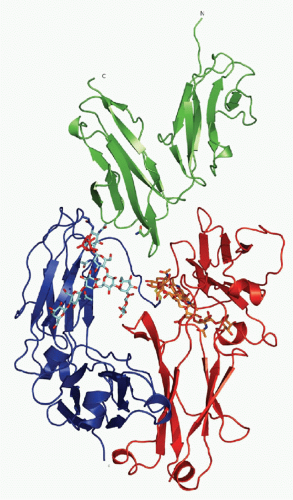 FIG. 24.4. Ribbon Diagram of Crystallizable Fragment (Fc) Receptor III-Immunoglobulin (IG)G1 Fc Structure. The carbohydrate moiety attached to both CH2 domains of the IgG Fc-fragment is shown as a stick and ball model. The extracellular domains of Fc receptor III are shown, together with the Fc fragment of IgG1. See text for details. Adapted from Sondermann et al.,15 with permission. |
The structure of the FcR bound to its ligand reveal that the antigen-binding fragment (Fab) arms are quite sharply bent and may adopt a perpendicular orientation toward the Fc. This arrangement would give the Fab arms maximal flexibility to bind antigen when the Fc fragment is oriented parallel to the membrane of the FcR-expressing cell. The asymmetrical interaction of the two Fc chains with a single FcR prevents a single antibody molecule from triggering dimerization of receptors and initiating signaling. Instead, dimerization is initiated by the interaction of antigen with the Fab arms, thus linking adaptive responses to effector cell triggering. Of note, the majority of the IgG-Fc&ggr;R cocrystals have been generated with Fc&ggr;Rs produced in bacteria and therefore in an aglycosylated form. In general glycosylation at Asn 297 is critical for Fc&ggr;R binding; modulation of Fc&ggr;R binding by glycan modification has been observed for several classes of glycan modifications, including sialylation, fucosylation, and branching N-acetyl glucosamine.85,86 The first IgG-Fc&ggr;R cocrystal structure with differentially glycosylated IgG variants demonstrated that carbohydrate-carbohydrate interactions between IgG and Fc&ggr;R are essential for high-affinity recognition of afucosylated IgG glycovariants by Fc&ggr;RIIIA.87 In contrast, low-affinity binding of Fc&ggr;RIIIA to fucosylated IgG is independent of these carbohydrate interactions. These novel insights may offer new ways of optimizing IgG-Fc&ggr;R interactions for enhanced therapeutic activity of IgG.
IN VITRO ACTIVITY
Binding Properties
As outlined in Figures 24.1 and 24.3, Ig binding to canonical FcRs falls into either high- or low-affinity binding classes. The high-affinity binding class is typified by Fc&egr;RI, with a binding affinity of 1010 M-1 for IgE, which ensures a monomeric interaction between IgE and its receptor. Fc&ggr;RI binds with relatively high affinity for IgG1 and IgG3 (human) and IgG2a (mouse) with an affinity constant of 108 M-1. In contrast to these high-affinity FcRs, the low-affinity receptors, such as the human Fc&ggr;RIIA, Fc&ggr;RIIB, Fc&ggr;RIIIA, Fc&ggr;RIIIB, and Fc&agr;RI, and the mouse Fc&ggr;RIIB, Fc&ggr;RIII, and Fc&ggr;RIV bind with affinities ranging from 5 × 105 M-1 (Fc&ggr;RIII) to 5 × 107 M-1. This low-affinity binding ensures that these receptors interact with ICs and not monomeric ligands. As described subsequently, this dependence on high-avidity and low-affinity interactions ensures that these receptors are activated only by physiologically relevant ICs and not by circulating monomeric Ig, thus avoiding inappropriate activation of effector responses. In general, low-affinity Fc&ggr;Rs bind IgG1 and IgG3 preferentially; their binding to IgG2 and IgG4 is of even lower affinity. As mentioned previously, polymorphisms in Fc&ggr;RIIA and Fc&ggr;RIIIA affect binding to IgG2 and IgG1, respectively, which may have significance in vivo in predicting responses to specific cytotoxic antibodies (Table 24.1). The isolated consideration of the affinity of antibody isotypes to their activating Fc&ggr;Rs is not sufficient, however, to explain the differences in in vivo activity. More importantly, the ratio of the affinities of an antibody isotype to the activating Fc&ggr;Rs compared to the inhibitory Fc&ggr;RIIB (termed A/I-ratio) predicts antibody activity more consistently.20,64 Thus, antibody isotypes with a high A/I-ratio, such as mouse IgG2a and IgG2b, will show a greater activity than counterparts with a lower ratio (see Table 24.1). The sugar moiety of the antibody Fc-portion is essential for FcR binding and the presence or absence of certain sugar residues can significantly impact on antibody-FcR binding.20,80,88,89 The absence of fucose, for example, will selectively increase the affinity of human IgG1 or mouse IgG2b for human Fc&ggr;RIIIa or mouse Fc&ggr;RIV, respectively. In contrast, the presence of terminal sialic acid on the N-linked Fc glycan reduces affinities for FcRs by an order of magnitude with a concomitant reduction in in vivo activity.80 Subunit interactions have also
been reported to influence affinity for ligand, as demonstrated for the common &ggr;chain associating with Fc&ggr;RIIIA.90 Its affinity for IgG1 is higher than the GPI-anchored form of this receptor, Fc&ggr;RIIIB.
been reported to influence affinity for ligand, as demonstrated for the common &ggr;chain associating with Fc&ggr;RIIIA.90 Its affinity for IgG1 is higher than the GPI-anchored form of this receptor, Fc&ggr;RIIIB.
TABLE 24.1 Affinities of Mouse and Human Fc Receptors to Different Antibody Isotypes | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||




