Biochemical Mediators of Allergic Reactions
Chhavi Gandhi
Stephen I. Wasserman
The cells and mediators involved in diseases of immediate-type hypersensitivity have been well described. The biologically active molecules responsible have been identified, and a thorough biochemical and structural elucidation of diverse lipid mediators has been accomplished. The activity of mediator-generating cells and their diverse products has been assigned a central role in both immunoglobulin E (IgE)-mediated acute and prolonged inflammatory events. More recently, a broadened understanding of the previously implicated mediators and newer agents has occurred both in human and animal model studies. This chapter places in perspective the mediator-generating cells, the mediators themselves and these newer concepts of their roles in pathobiologic and homeostatic events.
Mediator-Generating Cells
Mast cells and basophilic polymorphonuclear leukocytes (basophils) constitute the two cell types on which the high affinity FcεRI is expressed (1). Mast cells are most closely related to mononuclear leukocytes (2) and are richly distributed in the deeper region of the central nervous system, the upper and lower respiratory epithelium, the bronchial lumen, the gastrointestinal mucosa and submuscosa, bone marrow, and skin (3). They are especially prominent in bone, dense connective tissue adjacent to blood vessels (particularly small arterioles and venules)- and peripheral nerves. In the skin, lungs, and gastrointestinal tract, mast cell concentrations approximate 10,000 to 20,000 cells/mm3. (They develop from CD34+ bone marrow precursors through the action of stem cell factor (kit-ligand SCF), which binds to a specific receptor (c-kit, CD117) (4). Precursor cells exit the marrow and terminally differentiate in tissues under a variety of local influences, such as interleukin-3 (IL-3), IL-4, IL-6, IL-9, IL-10, and factors from fibroblasts (5).
Mast cells are large (10 to 15 mm in diameter) and possess a ruffled membrane, numerous membrane-bound granules (0.5 to 0.7 mm in diameter), mitochondria, a mononuclear nucleus, and scant rough endoplasmic reticulum. Ultrastructurally, human mast cell granules display whorl and scroll patterns. Mast cells are heterogeneous, and both connective tissue (MCTC) and mucosal types (MCT) have been recognized. MCTC dominate in the skin and can be distinguished from the mucosal type by expressing CD-88 (C5aR) on their cell surface. They are also found in lesser quantities in the gastrointestinal system, bronchial smooth muscle, and glandular region of the lungs. In the asthmatic lung, elevated levels of MCTC are found in the smooth muscle (6). This mast cell type expresses tryptase, chymase, cathepsin-G-like protease, and carboxypeptidase A in the granules. Mucosal-type mast cells (MCT) predominate in the lamina propria of the gastrointestinal tract as well as the peripheral airways, alveolar septa, and epithelium of the lung. MCT cells predominantly express tryptase (7) (Table 3.1). Recently, a c-kit mutant mouse has been developed using the W-sash mouse (Kit W-sh/W-sh) producing an experimental mast cell-deficient mice model that, unlike prior c-kit knockout models, can successfully reproduce (8), has no other leukocyte deficiencies, and should thus be helpful in further describing the mast cell’s role in inflammation.
Table 3.1 Human Mast Cell Subtypes | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Basophils, most closely related to eosinophils, are circulating leukocytes whose presence in tissue is unusual except in disease states. They originate in the bone marrow and constitute 0.1% to 2.0% of the peripheral blood leukocytes. Basophils possess a polylobed nucleus and differ from mast cells in their tinctorial properties, their relatively smooth cell surface, and their granule morphologic makeup, which is larger and less structured than that of the mast cell. Their growth is responsive, not to SCF, but rather to IL-3, and granulocyte-macrophage colony-stimulating factor (GM-CSF) (9).
Other mediator-generating cells include platelets, which express FcεRI and release serotonin, RANTES, platelet-activating factor (PAF), histamine, and platelet
factor 4, as well as cytokines and chemokines. Their role has been imputed from mouse studies of anaphylactoid shock in mast cell-deficient animals. Eosinophils have been suggested to play both a proinflammatory role, through release of granule-associated proteins, and an anti-inflammatory role, through the metabolism of vasoactive mediators (Table 3.1) (9).
factor 4, as well as cytokines and chemokines. Their role has been imputed from mouse studies of anaphylactoid shock in mast cell-deficient animals. Eosinophils have been suggested to play both a proinflammatory role, through release of granule-associated proteins, and an anti-inflammatory role, through the metabolism of vasoactive mediators (Table 3.1) (9).
Activation of Mast Cells and Basophils
Mast cells and basophils possess numerous high-affinity intramembranous receptors (FcεRI) for the Fc portion of IgE. The number of such receptors is upregulated and their stability enhanced by exposure of the mast cell or basophil to increased amounts of IgE (10). Aggregation of two or more of these FcεRI receptors with antigen cross-linking of receptor-bound IgE molecules leads to receptor activation and complex signal transduction leading to the release of mediators from mast cells and basophils (11). Preformed mediators from mast cell cytoplasmic granules are released immediately and include histamine, neutral proteases, a small proportion of total cytokines, and proteoglycans. Unstored mediator synthesis is initiated in minutes and includes the production of lipid membrane-derived arachidonic acid, prostaglandins (PGD2), PAF, and leukotrienes (LTB4 and LTC4). Chemokine and cytokine products are unstored and are produced within hours of the initial stimulus; their release can continue for days (12). Mast cell responsiveness may be heightened by exposure to SCF or other cytokines, whereas basophils are primed to respond by GM-CSF, IL-1, IL-3, and IL-5. Other important secreatagogues include a family of histamine-releasing factors and complement fragments C3a and C5a, the latter of which have not shown to be necessary to mount an anaphylactic episode. (The coagulation cascade and kallikrein-kinin contact system have also been implicated to be involved because of the reported decreases in fibrinogen, factor V and VIII, and factor XIIa-c1 inhibitor complexes, and the high-molecular weight kininogen and kallikrein-C1 that have been seen [13]).
The FcεRI receptor is composed of one extracellular subunit, α, which binds to IgE, and two intracellular subunits, β and γ, which are associated with enzymes and are essential in the subsequent signal transduction on the activation of mast cells and basophils. FcεRI receptor bridging is followed by phosphorylation of the immunoreceptor tyrosine-based activation motifs (ITAM) of the β and γ subunits, which acts as a scaffold to allow the binding of additional signaling molecules, the most important of which is the protein tyrosine kinase Syk. Syk binds, via the γ subunit of the receptor, becomes phosphorylated, and leads to the phosphorylation of several downstream proteins, directly and indirectly (11). The net result of this signaling is an increase in intracellular calcium, protein kinase C translocation, G protein activation, and cyclic adenosine monophosphate generation. At the same time, membrane phospholipids are metabolized to generate monoacylglycerols, diacylglycerols, and phosphorylated inositol species, which facilitate protein kinase C function and liberate Ca2+ from intracellular sites. While these biochemical events are underway, adenosine triphosphate (ATP) is catabolized and adenosine is liberated which further activates a mast cell adenosine receptor to enhance granule release. Finally, the cell gains control over mediator release, the process stops, and the cell regranulates (14).
Although initiated at the time of IgE and antigen activation, the generation of cytokines is expressed over a time frame of hours to days. Both mast cells and basophils are important sources of a variety of inflammatory cytokines, as described later. After the initiating event of allergen binding, cytokine synthesis proceeds through activation of such signaling pathways as the STAT and NF-κB-regulated processes, with gene transcription evident within hours and protein secretion occurring subsequently (15).
Recent work has added further complexity to mast cell and basophil activation and has suggested pathways for their regulation. Mast cells possess a receptor for IgG, FcγRII, which can modulate mediator release; these cells also respond to endotoxin through engagement of a Toll-like receptor complex. The presence of these additional modulatory pathways suggests that mast cell and basophil mediators participate in inflammatory conditions in which IgE may not be present. Murine models suggest further mechanisms in the activation and signaling of mast cells and basophils. Zinc has been implicated in inducing FCεRI-dependent mast cell degranulation, cytokine production (IL-6 and TNF-α), NF-κB activation, and possibly protein kinase C plasma membrane translocation (16). Sensory skin nerves may augment mast cell-derived inflammation by releasing neuropeptides such as substance P and calcitonin gene-related peptide in this setting, with less mast cell-driven inflammation being seen in denervated skin (17). A protein of the regulator of G-protein signaling (RGS) family, RGS13, has been shown to halt normal signal transduction downstream of FcεRI by blocking PIP3 phosphorylation from occurring in mast cells, suggesting it may have a role in their homeostasis (18). A human Ig fusion protein, Fcγ-Fcε (GE2 protein), has been shown to inhibit FcεRI signaling by cross-linking FcεRI with FcγRIIb, a negative regulatory molecule (19). Its role in the basal regulation mast cell and basophil activity has not been fully evaluated.
Mediators
Whatever their final metabolic interrelationships, the early biochemical processes lead to the generation of a heterogenous group of molecules termed mediators. Some mediators are preformed and are stored in the granules of the cell; others are generated only after cell activation and originate in the cytosol or membrane. Mediators are classified in this chapter by their proposed actions (Tables 3.2 and 3.3), although some mediators subserve several functions.
Table 3.2 Mast Cell Vasoactive and Spasmogenic Mediators | ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Table 3.3 Mast Cell Mediators Affecting Cell Migration | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||
Spasmogenic Mediators
Histamine, generated by decarboxylation of histidine, was the first mast cell mediator to be identified, and it is the sole preformed mediator in this functional class. It is bound to the proteoglycans of mast cell and basophil granules (5 and 1 mg/106 cells, respectively) (20). Histamine circulates at concentrations of about 300 pg/mL with a circadian maximum in the early morning hours. (Histamine excretion exceeds 10 mg/24 hours; a small fraction is excreted as the native molecule and the remainder as imidazole acetic acid or methyl histamine.) Histamine interacts with specific H1, H2, H3, and newly discovered H4 receptors (20). The H4 receptor has low homology with the other histamine receptors but shares the most with the H3 receptor with 35% amino acid homology; antihistamines specific for the H1 and H2 receptors do not bind to the H4 receptor (20).
All of the histamine receptors are G-protein coupled receptors. H1 receptors utilize Gq proteins, leading to phopholipase C activation, inositol phosphate production, and, eventually, calcium mobilization (20). H2 receptors use Gαs proteins, causing an increase in cyclic AMP. H3 receptors use Gαi/o, causing inhibition of cAMP, increasing calcium mobilization, and activating MAP kinases and ion channels. H4 receptors seem to work through pertussis-toxin sensitive Gαi/o proteins, which signal through increases in intracellular calcium; however, other pathways have been described (21).
H1 receptors predominate in the skin and smooth muscle; H2 receptors are the most prevalent in the skin, lungs, stomach, and on a variety of leukocytes; H3 receptors predominate in the brain; H4 receptors appear to be present on mast cells, basophils, eosinophils (22), dendritic cells, CD4+ effector T cells (at low levels)
(23), and may be present on neutrophils and monocytes (46) as well as on lung parenchymal cells (24). The biologic response to histamine reflects the ratio of these receptors in a given tissue. Increased levels of histamine have been reported in the blood or urine of patients with physical urticaria, anaphylaxis, systemic mastocytosis, and antigen-induced rhinitis and asthma (25). H1 histamine effects include the contraction of bronchial and gut musculature, vascular permeability, pulmonary vasoconstriction, and nasal mucus production (26). By its H2 pathway, histamine dilates respiratory musculature, enhances airway mucus production, inhibits basophils and skin (but not lung) mast cell degranulation, and activates suppressor T lymphocytes. Both H1 and H2 actions are required for the full expression of pruritus, cutaneous vasodilation, and cardiac irritability (20). The H3 actions of histamine suppress central nervous system histamine synthesis. The H4 actions of histamine include eosinophil chemotaxis, cell shape change, and up-regulation of adhesion molecules, the induction of mast cell chemotaxis toward a histamine gradient (22,23) and an increase in cytokine production from dendritic and T cells (24,27). Studies using the H4 receptor antagonist JNJ 7777120 help support these findings (28), and H4 KO mice suggest the H4 receptor has a role in allergic airway inflammation through activation of CD4+
 Evaluation and Management of Immune Deficiency in Allergy Practice
Evaluation and Management of Immune Deficiency in Allergy Practice
 Radiologic Evaluation of Allergic and Related Diseases of the Lower Airway
Radiologic Evaluation of Allergic and Related Diseases of the Lower Airway
 Drug Allergy Part A Introduction, Epidemiology, Classification of Adverse Reactions, Immunochemical Basis, Risk Factors, Evaluation of Patients with Suspected Drug Allergy, Patient Management Considerations
Drug Allergy Part A Introduction, Epidemiology, Classification of Adverse Reactions, Immunochemical Basis, Risk Factors, Evaluation of Patients with Suspected Drug Allergy, Patient Management Considerations
 Asthma
Asthma
 Nasal Polyposis, Sinusitis, and Nonallergic Rhinitis
Nasal Polyposis, Sinusitis, and Nonallergic Rhinitis
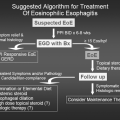 Eosinophilic Esophagitis
Eosinophilic Esophagitis




(23), and may be present on neutrophils and monocytes (46) as well as on lung parenchymal cells (24). The biologic response to histamine reflects the ratio of these receptors in a given tissue. Increased levels of histamine have been reported in the blood or urine of patients with physical urticaria, anaphylaxis, systemic mastocytosis, and antigen-induced rhinitis and asthma (25). H1 histamine effects include the contraction of bronchial and gut musculature, vascular permeability, pulmonary vasoconstriction, and nasal mucus production (26). By its H2 pathway, histamine dilates respiratory musculature, enhances airway mucus production, inhibits basophils and skin (but not lung) mast cell degranulation, and activates suppressor T lymphocytes. Both H1 and H2 actions are required for the full expression of pruritus, cutaneous vasodilation, and cardiac irritability (20). The H3 actions of histamine suppress central nervous system histamine synthesis. The H4 actions of histamine include eosinophil chemotaxis, cell shape change, and up-regulation of adhesion molecules, the induction of mast cell chemotaxis toward a histamine gradient (22,23) and an increase in cytokine production from dendritic and T cells (24,27). Studies using the H4 receptor antagonist JNJ 7777120 help support these findings (28), and H4 KO mice suggest the H4 receptor has a role in allergic airway inflammation through activation of CD4+
Related posts:
Evaluation and Management of Immune Deficiency in Allergy Practice
Radiologic Evaluation of Allergic and Related Diseases of the Lower Airway
Drug Allergy Part A Introduction, Epidemiology, Classification of Adverse Reactions, Immunochemical Basis, Risk Factors, Evaluation of Patients with Suspected Drug Allergy, Patient Management Considerations
Asthma
Nasal Polyposis, Sinusitis, and Nonallergic Rhinitis
Eosinophilic Esophagitis
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
