Animals born and reared in the complete absence of contact with any non-self material (not an easy procedure!) have virtually no immunoglobulins in their serum, but as soon as they encounter the normal environment, with its content of microorganisms, their serum immunoglobulin (Ig) rises towards the normal level of 10–20 mg (or about 60 000 000 000 000 000 molecules) per millilitre. This shows that immunoglobulins are produced only as a result of stimulation by foreign antigens, the process being known as the antibody response.
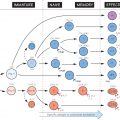
In the figure, these events are shown in a section through a stylized lymph node. Antigen is shown entering from the tissues (top left) and antibody being released into the blood (bottom right). The antigen is depicted as a combination of two components, representing the portion, or determinant, recognized by the B cell and against which antibody is eventually made (black circles) and other determinants that interact with T cells and are needed in order for the B cell to be fully triggered (white triangles). These are traditionally known as ‘haptenic’ and ‘carrier’ determinants, respectively. In practice, a virus, bacterium, etc. would carry numerous different haptenic and carrier determinants, whereas small molecules such as toxins may act as haptens only. But even small, well-defined antigenic determinants usually stimulate a heterogeneous population of B cells, each producing antibody of slightly different specificity and affinity.
The main stages of the response are recognition and processing of the antigen (see Fig. 18), selection of the appropriate individual B and T cells (shown larger in the figure), proliferation of these cells to form a clone, and differentiation into the mature functioning state. A prominent feature of all stages is the many interactions between cells, which are mediated mostly by cytokines (white arrows in the figure). There are also a number of regulatory influences whose relative importance is not yet clear. Most of these cell interactions occur in the lymph nodes or spleen, but antibody can be formed wherever there is lymphoid tissue.
In a subsequent response to the same antigen, average affinity tends to be higher, precursor T and B cells more numerous and Ig class more varied. This secondary response is therefore more rapid and effective, and such an individual is described as showing memory to the antigen in question; this, for example, is the aim of most vaccines (see Fig. 41).
AL
Afferent lymphatic, via which antigens and antigen-bearing cells enter the lymph node from the tissues (see Fig. 17).
APC
Antigen-presenting cell. Before they can trigger lymphocytes, antigens normally require to be presented on the surface of a specialized cell. These cells form a dense network within lymphoid tissue, through which lymphocytes move around searching for antigen. B cells recognize antigen on the surface of follicular dendritic cells (FDC), while T cells interact with interdigitating dendritic cells (IDC) which carry high levels of MHC and costimulatory
Related posts:


Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


