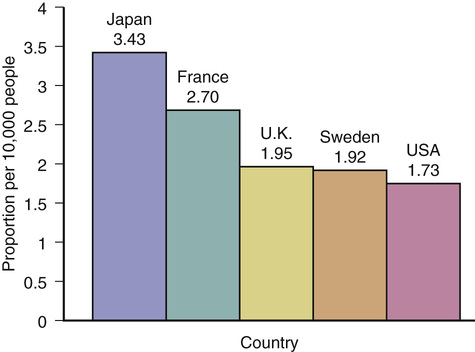
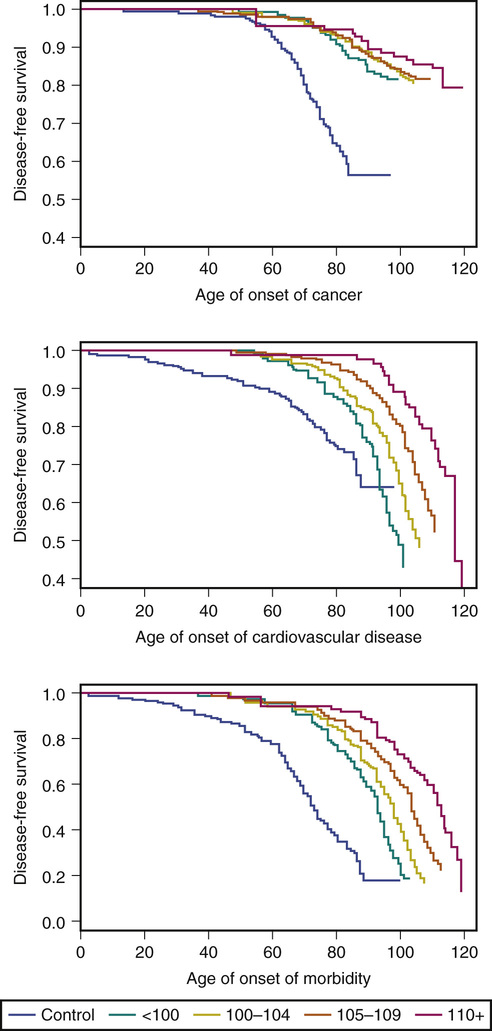
Thomas T. Perls According to the U.S. Social Security Administration, in 2010, approximately 51,000 people aged 100 years and older collected Social Security benefits.1 The U.S. census reported a similar number of 53,364 and an overall prevalence of 1.73 centenarians/10,000 people, with 80% of centenarians being women.1 In the 1980s and 1990s, centenarians were deemed the fastest growing age group in the population (65.8% from 1980 to 2000) but, in 2007, the Census Bureau’s Velkoff and Humes indicated that the earlier reported numbers were artificially too high.2 In its 2010 report on centenarians, the U.S. Census indicated a 5.8% increase in centenarians from 2000 to 2010, whereas the overall population grew by 9.7%. On the other hand, octogerians and nonagerians are the fastest growing groups, with 21% and 30% growth, respectively, over the same period of time. Figure 4-1 depicts the proportions of centenarians in other countries also noted by the census report on centenarians.3 It is impressive that the proportion in Japan is twice that of the United States. The oldest ever valid age claim is that of Jeanne Calment, who was from Southern France and died at the age of 122 years and 164 days in 1997.4 The record for a man was recently established by a Japanese man named Jiroemon Kimura, who died at the age of 115 years and 253 days in 2013 (birth date, April 19, 1897). It is not unusual to hear of claims of people exceeding these ages, but 99% of claims of ages older than 115 years are false.5 A clear tipoff that a claim is false is when someone is claimed to be the oldest person ever, and yet there was no mention of their age when they exceeded the current record of 122 years. For example, in 2009, the extraordinary age claim of Sakhan Dosova, of Kazakhstan, purported to be 130 years old (1879-2009), was published in a popular scientific journal, despite the fact that she never attracted attention when she surpassed 122 years and that there was no documentation supporting her being alive in the early 1880s.6 In 2014, according to the Gerontology Research Group (www.grg.org), there were approximately 62 supercentenarians (aged 110+ years) in the United States or a prevalence of about 1/5 million people. The Social Security Administration’s Kestenbaum and Ferguson counted 325 supercentenarians who died in the period 1980 to 2003 and 90% of these were female.7 In light of the above observations, the 2010 U.S. and Japanese census reports very likely list far too many supercentenarians, 330 (~1/400,000) and 711 (~1/180,000), respectively, speaking to the high false-positive rate for counts of supercentenarians in many national censuses.8,9 Although female centenarians outnumber their male counterparts by approximately 8 : 1, male centenarians tend to have significantly better functional status than their female counterparts. The fact that male centenarians more frequently have better physical and cognitive function has been noted in most centenarian studies, most notably the Italian Centenarian Study.10 A plausible hypothesis for why male centenarians fare better is that only those who are functionally independent are able to achieve such extreme old age. Women, on the other hand, appear to experience the double-edged sword of being able to live longer while also living more frequently with age-related illnesses and disability. This hypothesis is supported by a Danish study, in which 38% of men at age 98 years were functionally independent, but then this proportion rose to 53% among 100-year-olds. The proportion of women who were independent, however, continued to fall, from 30% of 98-year-olds to 28% of 100-year-olds.11 Another paradox is that although the male centenarians might be exceptionally fit relative to the women, they appear to have higher age-related, disease-associated mortality rates, so that once they do develop a disease, such as dementia or stroke, their mortality risk probably is much higher than it might be for women. Such hypotheses point to the possibility that women are much more resilient than men with regard to aging and age-related diseases. In the New England Centenarian Study (NECS; http://www.bumc.bu.edu/centenarian), centenarians and their family members were studied primarily because of our long-held belief that these individuals are a model of successful aging. By determining environmental and genetic factors that are more or less common compared to those of other groups of people, we should be able to determine risk factors for premature versus healthy aging and to formulate strategies that enhance a person’s ability to compress their disability toward the end of a longer life. In 1980, James Fries proposed his “Compression of Morbidity” hypothesis.12 This hypothesis states that as people approach the limit of their life span, they necessarily must compress the time that they experience diseases that affect mortality toward the end of their life. Previously, when the NECS investigated this hypothesis, with its sample of 424 centenarians, mean age 102 years, it was found that centenarians did not all exhibit this compression. Instead, a substantial proportion (43%), termed survivors, lived with at least one of 10 age-related diseases—heart disease, stroke, diabetes, cancer, dementia, chronic obstructive lung disease, osteoporosis, hypertension—for 20 years or more. Another 42%, termed delayers, lived with such a disease(s) between the ages of 80 and 99 years. Finally, those who had none of these diagnoses at the age of 100 years, or escapers, comprised 15% of the sample.13 Of note, a study of the oldest subjects in the Health and Retirement Survey found a similar proportion of escapers.14 Thus, our findings appeared to be inconsistent with the Compression of Morbidity hypothesis. On the other hand, it was also noted that on average, these subjects were disability-free until the age of 93 years.15 Thus there appeared to generally be a compression of disability among centenarians, even despite a substantial incidence of age-related morbidities. Somehow, it seems that people who survive to 100 and older deal with these age-related diseases more effectively than other people with such diseases who die at a younger age. The ability to deal with stressors and, more generally, age-related diseases, leads to the as yet poorly defined notions of adaptive capacity, functional reserve, and resilience, which may be important distinguishing features of the ability to achieve exceptional old age.16 We suspected that to observe the compression of the morbidity phenomenon, we needed to include subjects who truly survived near the limit of the human life span. There is a tremendous degree of selection (very large proportions of the sample die) that occurs between the ages of 100 to 104 years and 110+ years, and thus it would make sense that there could be a significant difference between these age groups in terms of determinants of survival. Thus, since 2007, we made a concerted effort to enroll and longitudinally follow as many people aged 105+ years as possible. With a total sample of 343 nonagerian siblings of centenarians, 884 100- to 104-year-olds, 430 105- to 109-year-olds, and 104 110+-year-olds, and 90% of the subjects deceased, we analyzed the ages of onset of cancer, cardiovascular disease, diabetes, dementia, and stroke.17 We found that the ages of onset of numerous diseases were increasingly delayed with the older and older ages of the subjects in our NECS sample. For example, in Figure 4-2, Kaplan-Meyer survival curves show this progressive delay in age of onset for cancer, cardiovascular disease, and overall morbidity, where at least one of the following became clinically apparent—cardiovascular disease, cancer, diabetes, dementia, and/or stroke. Consistent with the Compression of Morbidity hypothesis, controls (spouses of the offspring of centenarians or the offspring of parents with an average life expectancy) experienced a mean 17.9% of their lives with one or more age-related diseases, centenarians (100 to 104 years) with 9%, semisupercentenarians (105 to 109 years) with 8.9%, and supercentenarians with 5.2%. These findings have important implications for the study of the basic biology of aging. As Fries’ article indicated, the compression of morbidity toward the end of life would implicate an overall exhaustion of organ reserve as the cause of death in these individuals.12 Anecdotally, this is what we observed in most supercentenarians. Furthermore, this progressive rectangularization of the survival curve with older and older ages of death also suggests a limit to the human life span. Finally, the fact that most of the supercentenarians in our sample experienced morbidity and disability in only the last few years of their lives indicates substantial phenotypic homogeneity. This homogeneity suggests an increased power with these samples of oldest subjects to discover environmental and genetic determinants that they have in common that promote such exceptional survival. There do not appear to be specific health behaviors that are consistently associated with exceptional longevity. However, that is not to say that for many people achieving these extreme ages, certain behaviors such as smoking would have caused their death at a substantially younger age. Alternatively, in some groups, healthy habits such as a Mediterranean diet may be an essential ingredient for exceptional longevity. It would make sense in terms of evolutionary theory that different ethnicities, environments, and cultures lead to different combinations of genetic and environmental variants that predispose to exceptional survival. Some phenotypes have been associated with exceptional longevity, including certain personality types and maternal age. The NECS assessed personality traits in the children of centenarians and found that both the males and females scored below normative values for neuroticism and above normal for extraversion. For the other personality domains of the NEO Five Factor Inventory, the offspring scored within normal range.18 Similar findings were made with the offspring generation of the Long Life Family Study.19 These findings make sense in light of the substantial literature demonstrating associations between elevated neuroticism and morbidities such as hypertension and cardiovascular disease.20 Elevated extraversion could indicate a propensity to effectively establish social relationships conducive to better cognitive and psychological health. The Tokyo Centenarian Study, led by Hirose, found, however, that their centenarian subjects scored within the normal range for neuroticism but elevated openness, indicating the possibility that cultures and ethnicities can differ in regard to which personality traits are associated with exceptional longevity.21 As discussed later, there is a growing body of evidence for a substantial genetic influence on survival to the most extreme ages. An important question is what would be the selection pressure(s) for the evolution of longevity-associated genetic variants. The pressure to have a longer period of time during which women can bear children and therefore have more of them, with greater success in passing one’s genes down to subsequent generations, could be one such pressure.22 This hypothesis is consistent with the disposable soma theory, in which the tradeoff in energy allocation between reproductive fitness and repair and maintenance functions can be delayed when longevity-associated variants facilitate slower aging and the delay or prevention of age-related diseases that also adversely affect fertility.23 Several studies have noted an association between older maternal age and increased odds of exceptional survival. The NECS assessed maternal age history in its sample of female centenarians and a birth cohort–matched referent sample of women who survived to the cohort’s average life expectancy. Women who gave birth to a child after the age of 40 years (fertility assistance was not technologically available to this cohort) had four times greater odds of being a centenarian.24 Other studies have noted such a correlation.25–27 Numerous investigators are now searching for and investigating genes that influence reproductive fitness in terms of their ability also to influence rate of aging and susceptibility to age-related diseases.28–30 Early on in the NECS, we discovered several families in which multiple siblings lived to such extreme ages that the probability of such clustering being observed would be fewer than one family per all the families existing in the world at the time.31 Thus, for such families to exist, the family members with the trait must have some factors in common, and this could not be simply due to chance. Family members have genetic and nongenetic factors in common, and nongenetic factors have clearly been associated with increased probability of survival to older age, including education, socioeconomic status, access to health care, diet, environmental exposures (good and bad), refraining from tobacco use and excess alcohol, and others. Equally apparent is that some of these factors are not purely nongenetic because genetic associations have been noted for traits such as tobacco and alcohol use. Sibling studies allow for the estimation of sibling recurrent risk for a trait expressed by another sibling. An initial study by the NECS found that siblings of centenarians had about a four times increased relative risk of living to 100 years compared to birth cohort–matched controls who did not have parental exceptional longevity.32 That work was limited by a relatively small sample and the relatively young age of the centenarian probands. Similarly powered studies from Iceland and Utah have produced sibling relative risks in this range.33,34 Later, when the New England study had many more centenarians who also had reached a substantially older age, the sibling relative risk for males was 18 and, for females, 8.5.35 The increased relative risk noted in males has been observed in other studies and suggests that males are more dependent on a genetic component than females to achieve exceptional longevity.36 The heritability of aging, longevity, and life span has been noted by different Scandinavian twin studies and a study of Amish pedigrees to be approximately 25% to 30%. The ages at death of the subjects in these studies ranged from 73 (standard deviation [SD], 16) and 71 (SD, 17) years (52nd and 49th percentiles of survival).37,38 In a 1998 Swedish twins study of longevity, which also interchangeably uses the term life span, no men in the sample lived past the age of 89 years and about 2% of the female sample lived to the age of 90+ years. This study estimated the genetic component of longevity to be about 33%.39 In a 2001 study, “Heritability of Life Span in the Old Order Amish,” the authors investigated Amish pedigrees for parental and offspring ages of death for subjects born prior to 1890 who survived to at least the age of 30 years. The mean age of death was 71 ± 16 years, and about 7% of the sample was aged 90+ years, with only a few subjects aged 95+ years, and they estimated the heritability of both to be 25 ± 5%.40 With percentiles of survival nowhere approaching those of centenarians (e.g., <1 percentile), the previously mentioned twin and other studies have little to do with exceptional longevity, and research and review articles continue misguidedly to promote the idea that the heritability of survival to an extreme age does not change with older and older ages and is relatively low, at about 25%. Furthermore, many people erroneously equate heritability with genetic contribution. Instead, heritability is a measure of familiality, which is due to genetic and environmental factors that family members have in common and that can influence the trait in question.41 Consistent with these studies, the Seventh Day Adventist Health Study suggested that average humans can, in the setting of specific healthy behaviors, achieve an average life expectancy of 86 years.42 The NECS investigated sibling relative risk according to the proband’s percentile of survival and the year in which they were born. Analyzing survival data from 1917 sibships in which at least one sibling survived to the age of 90 years or older, we found that sibling relative risk increases with older and older age of the proband and with earlier birth year of the proband at these ages. The siblings of males surviving to the fifth percentile (age 90 for those born in 1900) have themselves a 1.7 times greater chance of living to the same age compared to population controls. Siblings of males and females surviving to the oldest 0.01 percentile of survival have a 35.6 times greater chance of surviving to the same age.43 It should be noted, however, that just because the chances are so much greater for these siblings than for the average person from the same birth cohort, it does not mean that they are assured of living to such an age because, after all, it remains an exceptionally rare phenotype. These findings also illustrate how very important it is for studies of the oldest old to describe precisely the phenotype they are studying by including the birth year cohort and the participants’ percentile(s) of survival.
Successful Aging
The Centenarians
Demography of Centenarians
Extraordinary Age Claims
The Gender Disparity
Successful Aging
Phenotypic Associations
Personality
Maternal Age
Familial Association and Heritability
Related posts:






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


