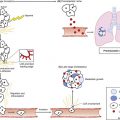
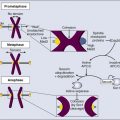
Piero Dalerba, Michael F. Clarke, Irving L. Weissman and Maximilian Diehn • Many tumors originate in organs and tissues that undergo a continuous process of cell turnover, which is sustained by a minority population of stem cells (e.g., the colon, breast, lung, prostate, brain, and bone marrow). • Stem cells have four fundamental properties: the ability to give rise to new stem cells with intact and unlimited growth potential (self-renewal), the ability to give rise to a progeny of specialized cells (differentiation), the ability to migrate into new tissue locations and establish tissue growth (migration and tissue repair), and the ability to balance the previous three properties according to a genetic program that places constraints on their numerical expansion (homeostatic control). • In many tissues, stem cells are the only long-lived cells. This observation suggests that early transforming events, either genetic mutations or epigenetic modifications, are likely to accumulate in stem cells. • In addition to oncogenes that control cell survival and proliferation, there is a class of oncogenes that regulate self-renewal. In some cancers, tumor growth might be sustained by progenitor cells, which do not naturally self-renew but have aberrantly acquired this ability during disease progression. • Experimental data suggest that, in many leukemias and solid tumors (e.g., breast, colon, head and neck, pancreas, bladder, and prostate carcinomas), only a specific, phenotypically distinct subset of cancer cells is able to form tumors when transplanted in mice. • Currently, most antitumor drugs are evaluated on the basis of their capacity to rapidly reduce tumor size. However, because self-renewing cancer cells are often minority populations, treatments that simply reduce tumor size may not effectively target these cells. • To result in cure, therapies must eradicate self-renewing cancer cells. The ability to identify these cells should allow the identification of new diagnostic markers and therapeutic targets. Many tumors originate from tissues that naturally undergo a continuous process of cell turnover. In such tissues, cell maturation is arranged according to a hierarchical system in which a minority population of stem cells is able to perpetuate itself through a process called “self-renewal” while also giving rise to several stages of intermediate progenitors and, eventually, to terminally differentiated cells (Fig. 7-1). As opposed to stem cells, intermediate progenitors and terminally differentiated cells have a limited expansion potential and are unable to self-renew.1,2 Because stem cells are frequently found in small numbers, they must be isolated and tested prospectively to define their molecular and biochemical properties. Although most tissues are likely to regenerate from tissue-specific stem cells, stem cells have been identified in only a few of them. The stem cells that give rise to the lymphohematopoietic system, called hematopoietic stem cells (HSCs), are among the best characterized and have been isolated from mice and humans.3–6 HSCs have important applications in cancer therapy, where bone marrow transplantation is used to regenerate the hematopoietic system after myeloablative treatments.6 The ability to isolate HSCs with high degrees of purity enables the execution of tumor-free autologous bone-marrow transplants in cancer patients.7,8 Understanding the biology of the normal tissues from which tumors originate, and especially of their stem cell populations, can provide important insights into cancer biology. Several aspects of stem cell biology are relevant to cancer.9,10 First, many cancer cells share with normal stem cells the capacity to self-renew, and emerging evidence suggests that similar molecular pathways underpin this property in the two cell types.11–16 Second, because stem cells are long-lived and often represent the only long-lived cells of normal adult tissues, they also represent the most likely target population where transforming events, either genetic mutations or epigenetic modifications, progressively accumulate, at least in the initial phases of the disease.2 Third, increasing evidence suggests that many tumor tissues, akin to normal ones, contain a minority “cancer stem cell” (CSC) population with indefinite proliferative potential that drives tumor growth and metastasis.17–20 Finally, many physiological properties of stem cells are mirrored in cancer cells and help define their biology. For example, because stem cells are long-lived and are necessary to maintain tissue homeostasis over the lifetime of an organism, they are frequently endowed with high levels of enzymatic systems that protect them from environmental hazards. These very same enzymatic systems endow specific subsets of cancer cells with resistance to antitumor agents and might help explain treatment failure. HSCs are the most studied and best understood type of somatic stem cells and serve as a model for stem cells in other tissues.21,22 Hematopoiesis is a tightly regulated process in which a small pool of HSCs gives rise to an increasingly diversified population of oligo-lineage intermediates, which in turn form the full repertoire of mature elements of the lymphohematopoietic system (e.g., erythrocytes, platelets, granulocytes, macrophages, and lymphocytes; Fig. 7-1).23,24 These mature cell types perform a diverse set of functions, ensuring tissue oxygenation, blood coagulation, and immunity. HSCs have four fundamental properties: self-renewal, differentiation, migration, and homeostatic control. First, HSCs need to self-renew to maintain the stem cell pool. Self-renewal is not synonymous with proliferation. Self-renewal is a cell division in which one or both of the daughter cells remain undifferentiated and maintain the identity of a stem cell, endowed with unlimited growth potential. Second, HSCs must undergo differentiation: they must generate a progeny of cells that expand and progressively specialize in order to replenish the various populations of mature cells that are damaged, aged, or otherwise lost. Third, HSCs are able to enter the circulation and migrate from one blood-forming site to another (e.g., from the marrow of one bone to that of another) to replace HSCs that have been lost and to maintain a constant output of blood cells.25 Finally, HSCs must balance self-renewal, differentiation, migration, and tissue repair according to a strict genetic program, which ensures homeostatic control of their numbers and regulates their expansion in response to various types of stress, such as bleeding or infection.28–28 In mice, HSCs represent less than 0.05% of bone marrow cells and are admixed with a heterogeneous pool of other immature progenitors. This pool includes a specific subset of immature multipotent progenitors (MPPs), which are able to differentiate into the full variety of hematopoietic lineages but lack self-renewal capacity.29 These populations form a hierarchy in which HSCs give rise to MPPs, which in turn give rise to oligo-lineage progenitors and the various mature cell types of the blood and lymphoid tissues (Fig. 7-1).6,24 As HSCs mature to become MPPs, they increase their proliferation rate but lose the ability to self-renew. Only HSCs can reconstitute hematopoiesis in the long term, for the lifetime of the animal, whereas MPPs can reconstitute hematopoiesis for a short period, usually less than 16 weeks.30 A similar hierarchy is also mirrored in human blood tissues, in which HSCs and MPPs are characterized by a CD34+CD38− phenotype and by the lack of mature blood cell lineage markers (Lin−) but can be separated on the basis of the differential expression of CD90 in HSCs (Fig. 7-1).31 More generally, the hierarchical organization of hematopoietic tissues can be successfully applied to model the cellular composition and developmental dynamics of many, if not most, adult mammalian tissues, such as the brain, mammary gland, and intestinal epithelium.32–35 The long-term survival of a tissue, either normal or neoplastic, is dependent on its capacity to self-renew, whereas its overall size is determined by the balance between the rates of cell proliferation and cell death across its various components.36 In normal tissues, stem cell numbers are under tight genetic regulation, resulting in the long-term maintenance of a constant tissue size.28–28 In contrast, tumor tissues have escaped this homeostatic regulation. Within a tumor, the number of cells with the ability to self-renew is constantly expanding, resulting in continuous tissue growth. Thus it is not surprising that many known oncogenes are able to expand stem cell numbers, either by protecting them from apoptosis or by inhibiting their differentiation. For example, enforced expression of Bcl2 results in an expansion of HSCs,37 whereas enforced expression of c-myb and c-myc prevents HSC differentiation along the erythroid lineage.38,39 Because the size of both normal and cancer tissues is dependent of the number of cells able to self-renew, it is also not surprising that a specific subset of oncogenes and/or tumor-suppressor genes might directly activate and/or disable self-renewal pathways.9,40 Indeed, the basic molecular machinery that ensures the unlimited growth capacity of most cancer cells (e.g., the telomerase enzymatic complex) is fundamental to ensure self-renewal of most normal stem cells.13–13 The capacity to modulate self-renewal, however, is not a necessary attribute of all oncogenes and/or tumor-suppressor genes; enforced expression of Bcl2, for instance, although able to expand the number of self-renewing HSCs, is unable per se to endow self-renewal properties on more mature cell types, such as MPPs.37 Among cancer genes with direct control over self-renewal functions, the best example is probably the Bmi1 oncogene. Bmi1 is a member of the Polycomb gene family, a group of transcriptional repressors involved in the epigenetic regulation of developmental and differentiation processes. In mice, Bmi1 can cooperate with c-myc to induce lymphoma.41 In mice genetically deficient for Bmi1 (Bmi1−/−), the number of HSCs is markedly reduced at birth, and their transplantation in lethally irradiated animals is able to reconstitute hematopoiesis, but only in a transient and self-limiting fashion, indicating a cell autonomous defect of self-renewal.14 Bmi1 exerts similar roles across many types of adult stem cells, such as neural and mammary stem cells.42,43 Lack of Bmi1 expression is associated with upregulation of tumor suppressors, such as the cyclin-dependent kinase inhibitors encoded by the alternative transcripts of the Cdkn2a locus (i.e. the p16Ink4a and p19Arf proteins), and with overexpression of transcription factors that promote differentiation, such as selected members of the Hox gene family.14 This association suggests that, in stem cells, the function of Bmi1 is to prevent the expression of a cascade of genes whose coordinated activity can lead to loss of self-renewal capacity. Remarkably, in a mouse model of leukemia initiated in Bmi1−/− mice, leukemic cells are initially able to expand and cause the death of their primary hosts but are unable to sustain long-term growth when transplanted in secondary syngeneic animals, where they eventually arrest, differentiate, and undergo apoptosis.15 Infection of Bmi1−/− leukemic cells with a retrovirus encoding for Bmi1 completely rescues this defect, allowing leukemic cells to grow indefinitely and be serially passaged in mice.15 Similar observations can also be obtained in human breast cancer, where downregulation of BMI1 by means of enforced expression of microRNA-200c associates with reduced expansion and tumorigenicity of cancer cells.16 Once more, this defect can be rescued by enforced expression of a modified version of the BMI1 messenger RNA, which is insensitive to the inhibitory action of microRNA-200c.16 Interestingly, human BMI1 appears to induce telomerase activity.44 Again, in many human model systems, cancer cells lacking telomerase, although capable of substantial short-term proliferation and numerical expansion, progressively exhaust their growth capacity and irreversibly arrest.13 These studies reinforce the concept that, in order to endow cancer cells with unlimited growth potential—a property frequently referred to as “immortality”36—constitutive activation of proliferation pathways is not sufficient. It is also necessary to ensure activation of self-renewal pathways and/or inactivation of pathways that prevent self-renewal. The Wnt pathway was first implicated in a mouse model of breast cancer, in which aberrant expression of Wnt1, caused by insertion of the mouse mammary tumor virus (MMTV) close to the Wnt1 gene, resulted in mammary tumors.45,46 Wnt1 belongs to a large family of secreted proteins that bind to receptors of both the Frizzled and low-density lipoprotein receptor–related protein (Lrp) families, resulting in activation of β-catenin.47 In the absence of Wnt stimulation, β-catenin is degraded by the adenomatous polyposis coli (Apc), glycogen synthase kinase–3β (Gsk3β), and Axin protein complex.48 Wnt/β-catenin signaling plays a pivotal role in the self-renewal of normal stem cells.49 Activation of β-catenin signaling by Wnt proteins allows expansion of stem/progenitor cells, both in vitro and in vivo, and across different tissues, including the bone marrow, skin, mammary gland, and small intestinal epithelium.50–53 Constitutive activation of the β-catenin pathway is oncogenic54,55 and is almost invariably observed in colon cancer, most frequently by inactivating mutations of human APC.56 Current evidence suggests that β-catenin signaling is key for cancer cells to maintain a stem/progenitor cell phenotype.19,57 In colon cancer cells, inhibition of the β-catenin pathway induces expression of p21cip1/waf1, a cell-cycle inhibitor, followed by proliferation arrest and acquisition of a differentiated phenotype.57 Enforced expression of c-myc, an oncogene whose transcription is activated by β-catenin, inhibits p21cip1/waf1 expression and allows cancer cells to continue proliferating in the absence of β-catenin signaling, revealing yet another role for a classic oncogene, c-myc, in the regulation of cell differentiation.57 The Notch family of receptors, first identified as regulators of wing patterning in the Drosophila fruit fly, control development and differentiation in many tissues and across many animal species.58 In vitro stimulation of HSCs with selected Notch ligands (i.e., Jagged-1 and Delta) transiently increases the activity of stem/progenitor cells, both in vitro and in vivo, suggesting that Notch activation promotes expansion of either HSCs or MPPs.59,60 Constitutive activation of the Notch pathway is endowed with powerful oncogenic effects in both mouse and human cells. The mouse oncogene int-3 encodes a constitutively active, truncated variant of the Notch-4 receptor.61 In humans, aberrant activation of NOTCH1, either by point-mutation or chromosomal translocation, is a common occurrence in T-cell acute lymphoblastic leukemia (T-ALL).62,63 Importantly, inhibition of NOTCH1 signaling can induce apoptosis in T-ALL cell lines and is now being explored as a promising therapeutic strategy.64 The Shh pathway provides yet another example of a pathway with key roles in both tissue development, stem-cell homeostasis, and oncogenesis. Like the Wnt factors, Shh is a secreted molecule and a powerful morphogen.65 Shh acts as a ligand for receptors encoded by members of the Patched gene family65 and activates signaling circuitries that regulate self-renewal in selected types of stem cells, such as bladder stem cells.66 Germline mutations in the SHH gene cause aberrations of embryo development in Drosophila and holoprosencephaly in humans.40 With relation to human cancer, germline mutations in Patched-1 (PTCH1) cause Gorlin or basal cell nevus syndrome (BCNS), whereas sporadic mutations in PTCH1 are observed in the majority of sporadic basal cell carcinomas (BCC) and a substantial fraction of medulloblastomas.40 Most importantly, pharmacological inhibition of the SHH pathway has recently shown powerful antitumor activity in clinical trials, against both human BCCs and medulloblastomas harboring PTCH1 mutations, and is rapidly changing treatment guidelines for these diseases.67–70 If oncogenic mutations often target signaling pathways that regulate self-renewal, then are stem cells the targets of neoplastic transformation? Several lines of evidence suggest that stem cells might be involved in the initial phases of cancer development. First, the fact that multiple and sequential mutations are necessary for a cell to become cancerous71,72 suggests that, in many cases, mutations might need, at first, to accumulate in a stem cell. Because mature cells often have a limited life span, it is unlikely that the full repertoire of mutations necessary to achieve malignant transformation might occur during their relatively short life. Second, many tumors arise in tissues that are known to contain stem cell populations, which intrinsically are endowed with the ability to self-renew. Because cancer cells must undergo self-renewal to achieve unlimited growth, stem cells might be more easily poised to undergo early malignant transformation steps compared with more mature cells, which naturally lack this fundamental property and would need to aberrantly activate self-renewal pathways to become malignant. However, even if the first set of initiating events is likely to target a stem cell population, it is possible that, as part of disease progression, the aberrant progeny of mutated stem cells might acquire additional changes and eventually gain the capacity to self-renew. In this scenario, the malignant clone would acquire a second population of self-renewing cells, whose phenotypic identity might resemble that of a progenitor rather than a stem cell (Fig. 7-2).2 Experimental evidence in support of a stem cell origin of cancer, at least during early initiation phases, is provided by elegant experiments in a mouse model of glioblastoma, caused by conditional deletion of the Neurofibromatosis-1 (Nf1), Trp53, and Pten tumor-suppressor genes.73 In this model, combined deletion of the three genes in brain tissues leads to rapid development of high-grade gliomas with 100% penetrance. This effect, however, can be observed only when the deletion is specifically targeted to cell compartments that include stem cell populations, either by conditional knockout of the tumor-suppressor genes in cells expressing the neural stem/progenitor marker Nestin or by somatic ablation of the tumor-suppressor genes in the mouse brain’s subventricular zone, where neural stem cells reside, which is achieved by stereotactic injection of an adenovirus encoding the Cre recombinase in mice genetically engineered to carry Nf1, Trp53, and Pten alleles between loxP sites. Most remarkably, somatic ablation of the same tumor suppressors in other areas of the brain, such as the striatum, is unable to generate any tumor. Moreover, tumors generated from adenovirus injection in the subventricular zone are able to disseminate throughout the brain parenchyma, following natural migration pathways of neural stem cells and exhibiting phenotypic hallmarks of multilineage differentiation along the astrocytic, neuronal, and oligodendrocytic lineages. Although it is likely that a first set of “early” transforming mutations might accumulate in the stem cell compartment, it is also possible that more mature, downstream progenitors that originated as the progeny of mutated stem cells might acquire a second set of “late” mutations, which might constitute the ultimate transforming event giving rise to cancer.74 In this scenario, mutated stem cells might represent a “reservoir” population of precancerous cells, whereas fully transformed progenitors might sustain the growth of the full-blown neoplastic mass (Fig. 7-2).75 This concept is supported by studies on normal hematopoietic development and hematologic malignancies, where the developmental hierarchy of the different cellular compartments is best understood, and where phenotypic differences between HSCs and MPPs allow a careful dissection of their relative contributions. For example, in mice, simultaneous deletion of the Trp53 gene and the Cdkn2a locus (encoding the p16Ink4a and p19Arf tumor suppressor proteins) leads to abnormal and selective acquisition of self-renewal properties by MPPs, which remain capable of producing the various differentiated cell types, but also acquire self-renewal and become capable of supporting the production of blood tissues upon serial transplantation, in essence behaving as stem cells.76 This observation indicates that genetic mutations commonly observed in human cancer might be responsible for a progressive and pathological “unleashing” of self-renewal properties in cells that are usually not endowed with them. Similar observations have also been made for different forms of human leukemia, such as acute myelogenous leukemia (AML) and chronic myelogenous leukemia (CML). One of the most frequent mutations in AML is the t(8;21) translocation, which results in the expression of a chimeric AML1/ETO transcript.77 Marrow samples from patients with t(8;21) AML treated at Hiroshima Hospital, when collected at clinical remission, contained HSCs (CD34+CD90+CD38−Lin−) that had up to 90% incidence of the chimeric AML1/ETO transcript.78 When these mutated HSCs were analyzed in vitro for their differentiation capacity, they gave rise to normal myeloerythroid progenies, showing that the mutation, alone and by itself, was insufficient to confer a fully transformed phenotype. Remarkably, marrow samples from the same patients, when collected at relapse, contained high numbers of cells with an MPP phenotype (CD34+CD90−CD38−Lin−) that gave rise to leukemic “blast” colonies in vitro.78 This observation is common to many forms of AML79 and is associated with the expansion of a pathological population of MPPs that have abnormally gained the capacity to self-renew.80 Taken together, these observations suggest that, in human AML, a first set of mutations (e.g., AML1/ETO) might initially accumulate in stem cells and that subsequent mutations in either the stem cells or their progeny might lead to overt leukemia.2 A similar scenario is observed in patients with CML, where leukemic cells are characterized by the t(9;22) BCR/ABL chromosomal translocation. In the initial “chronic phase” of human CML, the leukemic population undergoes a multilineage differentiation process, similar to that sustained by normal HSCs.81,82 At this initial stage, β-catenin signaling is active in HSCs but not in their descendant progenitor cells. However, when CML progresses to “myeloid blast crisis,” patients experience an AML-like disease. In this second stage, the leukemic clone acquires a second, phenotypically distinct, self-renewing population whose surface marker profile corresponds to that of a granulocyte-macrophage progenitor cell (GMP) and that is able to transplant the disease to immunodeficient mice.83 This second population is characterized by de novo aberrant activation of the β-catenin pathway, often as a result of a pathological mis-splicing of the GSK3β inhibitory enzyme.84
Stem Cells, Cell Differentiation, and Cancer
Introduction
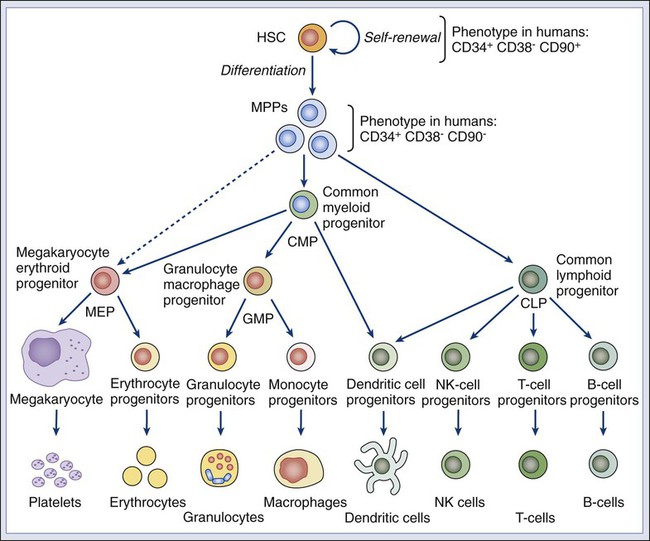
Properties of Normal Stem Cells
Genetic Regulation of Self-Renewal
Target Cells for Malignant Transformation
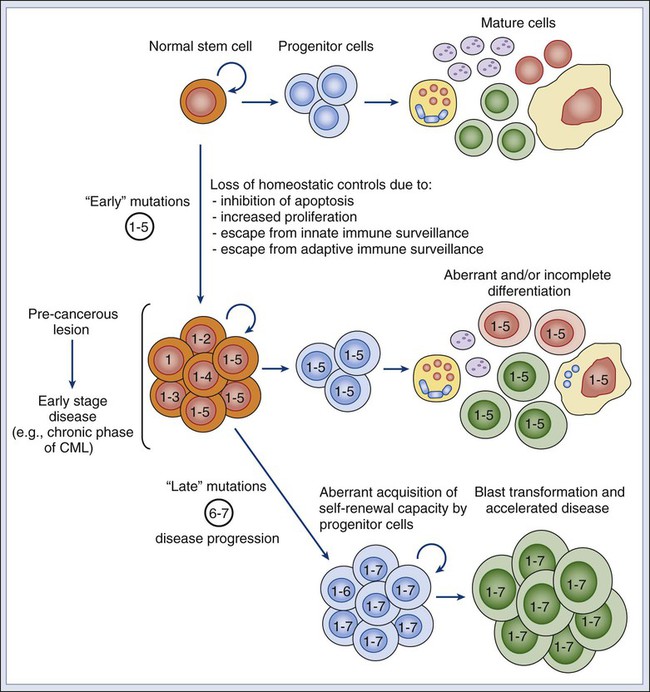
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Oncohema Key
Fastest Oncology & Hematology Insight Engine