Fig. 4.1
Normal distribution of inflammatory cells at the feto-maternal interface. Decidual natural killer and macrophages accumulate near the maternal spiral artery during the remodeling process. The vessel undergoes dramatic anatomic modification, lacking the smooth muscular wall and the elastic fibers and transforming in a low-pressure, high-conductance vessel. Inflammatory cells play a role in the removal of dying cells and cellular debris, to facilitate trophoblast migration and invasion. Moreover, inflammatory cells contribute to maternal tolerance versus the semi-allograft conceptus. Extravillous trophoblasts invade into the decidua from the anchoring villi and progressively replace the endothelium of the vessel. This type of extravillous trophoblast takes the name of “endovascular trophoblast,” and it is directly in contact with the maternal circulation. To avoid maternal reject reaction, extravillous trophoblast expresses a nonclassical major histocompatibility complex class I: it expresses a specific type of human leukocyte antigen (HLA), the HLA-G. HLA-G enters the maternal circulation and binds with the leukocyte immunoglobulin-like receptor on decidual natural killer cells, macrophages, and T lymphocytes, inactivating leukocytes and, in turn, inducing tolerance
The second trimester of pregnancy is a phase of predominant anti-inflammatory state. In this time frame, the mother and fetus are in symbiosis and the fetus undergoes rapid development and growth [5].
The late third trimester of pregnancy is a phase characterized by a reconversion to a pro-inflammatory condition that characterizes human parturition of an altricial newborn [5].
4.2 Natural Killer Cells
In pregnancy, decidual natural killer cells have a characteristic and specific phenotype (CD56BRIGHT, CD16−, CD3−). They change their structure as pregnancy progresses to adapt themselves to the role of pregnancy supporters.
During the first trimester, NK cells contain specific granules rich in angiogenic growth factor and vascular endothelial growth factor C. Moreover, during the first stage of pregnancy, decidual NK cells secrete matrix metalloproteinases MMP2 and MMP9. These enzymes break down the extracellular matrix. This process directly helps trophoblast migration and invasion. At this stage, NK cells play a decisive role for the proper placental development, promoting spiral artery remodeling. The number and the function of uterine NK cells are crucial for the physiological evolution of pregnancy. Their abnormalities have been associated with implantation failure [8], miscarriage [9], preeclampsia, and fetal growth restriction [10].
In the second trimester, decidual NK cells undergo a degranulation process. This will stop the production of angiogenic and vascular growth factors. Interferon-γ (INF- γ) secretion starts to oppose vascular development. During the third trimester, the uterine NK cells in the decidua are degranulated. In this stage, their production of INF-γ inhibits the trophoblastic invasiveness, protecting the uterus from a too deep trophoblastic migration [6].
4.3 Macrophages
At the feto-maternal interface, macrophages represent the second most abundant leukocyte population within the human decidua. Commonly, macrophages are classified in two different types: M1, the proinflammatory type, and M2, the immune-regulatory type [11]. The type of decidual macrophages appears to be predominantly M2 [12]. Their function is that of immune suppression, scavenging of apoptotic cells, clearing cellular debris, and promoting tissue remodeling.
4.3.1 Decidual Macrophages
Decidual macrophages are maternal cells and protect human pregnancy throughout gestation by inhibiting the immune response by decidual NK, T lymphocytes, and other circulatory macrophages [6]. Thanks to their additional production of matrix metalloproteinases 9 and vascular endothelial growth factor, decidual macrophages are believed to play a role in the spiral artery remodeling [12]. Despite this role in spiral artery modification, the association between abnormal number or function of decidual macrophages and preeclampsia is controversial [7].
4.3.2 Placental Macrophages
Placental macrophages, also called “Hofbauer cells,” are CD68+ fetal cells early detectable in pregnancy in the villous stroma. Since Hofbauer cells appear in the placental villi prior to the appearance of the fetal circulation, they may originate in the early stage of pregnancy from progenitor cells within the population of mesenchymal cells in the villous stroma; then, later in pregnancy, they may originate from penetration of embryonic bone marrow-derived monocytes into the villous stroma [3]. Generally, macrophages intervene in phagocytosis of apoptotic bodies and cellular debris as well as antigen presentation in response to inflammation and infectious insults; the specific role of placental macrophages has not been fully understood yet [13]. Their close contact with endothelial progenitor cells and primitive vessels suggests that Hofbauer cells intervene in early placental vascular genesis [14]. Moreover, the identification of proteins regulating branching morphogenesis localized to Hofbauer cells permits to hypothesize a role of placental macrophages in the development of the placental villous tree [15]. Alterations in the number and aspect of Hofbauer cells may be associated with complications of pregnancy such as fetal immune or nonimmune hydrops and fetal metabolic storage diseases such as GM1 gangliosidosis, glucuronidase deficiency, and type VII mucopolysaccharidosis [13]. Histological features of elevated concentration of Hofbauer cells have been related to adverse pregnancy outcome in association with villitis of unknown etiology or acute chorioamnionitis (see below) [13].
4.4 T Cells
T cells are poorly represented within the human decidua. Their role is not completely understood.
During the early stages of pregnancy, a successful implantation occurs in a finely regulated pro-inflammatory/anti-inflammatory microenvironment; T cells seem to play a crucial role in this balance. T-helper (Th) cells can be classified into Th1 cells, which are involved in cellular immunity, and Th2 cells, which are involved in humoral immunity [16]. In the past, maternal tolerance of the semi-allogenic graft was explained by an absolute predominance of the Th2-type immunity, which overrules Th1-type immunity, therefore protecting the fetus from maternal Th1-cell attack [17]. More recently, the role of Th1 has been reconsidered, showing that Th1 activity plays an important role in pregnancy, promoting Th2 response, regulating the placentation process, defending the maternal–fetal microenvironment against infections, and later cooperating in the initiation of delivery [18]; therefore, the concept of absolute Th2 predominance has been reconsidered and the hypothesis of “Th1–Th2 cooperation” has been introduced [18]. Moreover, it has been suggested that differences in the expression of the Th1 pro-inflammatory action and Th2 anti-inflammatory action depend on the phase of pregnancy: early and late pregnancy needs a Th1 milieu, whereas Th2 cells prevail in mid gestation (Table 4.1) [5]. Abnormal pregnancy outcome may occur in the presence of an aberrant Th1/Th2 profile: an excessive Th1 reaction may be associated with recurrent miscarriage, preterm delivery, and preeclampsia [18, 19], but even an increase of Th2 secretion at the feto-maternal interface could be associated with miscarriage and preeclampsia [18].
Table 4.1
Inflammatory balance in pregnancy
Phase of pregnancy | Predominant immunological features | Physiological events | Comment |
|---|---|---|---|
Early pregnancy | Th1 | Blastocyst apposition to the uterus | In this phase, an inflammatory microenvironment needs for removing of apoptotic cells and cellular debris, for achieving an adequate repair of the uterine epithelium, and for the maternal tolerance versus the semi-allogenic embryo |
Decidual breaking for the blastocyst implantation | |||
Trophoblast invasion of the decidua | |||
Breaking into the spiral artery and disruption of the smooth muscular wall by the extravillous trophoblast, to warrant an adequate placental–fetal blood supply | |||
Trophoblast replacement of the spiral arteries endothelium | |||
Mild pregnancy | Th2 | Placental development | In this phase, an anti-inflammatory status predominates, but the mother maintains her ability to respond against infective–immunological insults |
Fetal growth | |||
Fetal organ maturation | |||
Late pregnancy | Th1 | Cervical ripening | In this phase, a recrudescence of an inflammatory status is necessary to promote parturition |
Myometrial contraction | |||
Labor and delivery | |||
Expulsion of the placenta | |||
Uterine involution |
Histological features of placental elevated concentration of T cells have been associated with adverse pregnancy outcome as may occur in chronic deciduitis, chronic chorioamnionitis, and villitis of unknown etiology [7] (see below).
4.4.1 T Regulatory Cells
T regulatory cells (Treg) are a subset of T lymphocytes that act as strong suppressors of inflammatory immune response with a fundamental role in preventing destructive immune response and auto-inflammatory and autoimmune disease, warranting peripheral self-tolerance and immune homeostasis [20, 21]. Treg cells levels are high in maternal blood during the first and second trimesters of pregnancy and decline prior to delivery and postpartum [20]. Moreover, Treg cells physiologically accumulate in the maternal decidua. A deficiency in Treg cells number has been associated with recurrent miscarriage and preeclampsia [20]. Indeed, it is of interest to underline that there are two developmental pathways of Tregs: thymic (tTreg) and extrathymic or peripheral (pTreg). tTregs appear to suppress autoimmunity, whereas pTregs operate to downregulate responses to antigens, such as those from ingested food, symbiotic bacteria, and allergens. Genomic analysis allowed to hypothesize that pTregs emerged in eutherian (placental) mammals to protect from maternal–fetal conflict. As a matter of fact, conserved noncoding sequence 1 (CNS1), which is required for pTreg development, has been detected present in all placental mammals, whereas it is absent in non-placental mammals such as the wallaby, opossum, and platypus, as it is expected to be absent in non-mammals such as zebra fish. pTregs specifically sort out paternal antigens in the trophoblastic cells and suppress effector T cells in an active process to induce tolerance [22]. Figure 4.2 shows the quasi-perfect correspondence between wild-type pregnant mice and knocked-out ones as regards embryonic reabsorption and number of females that presents reabsorbed embryos [23].
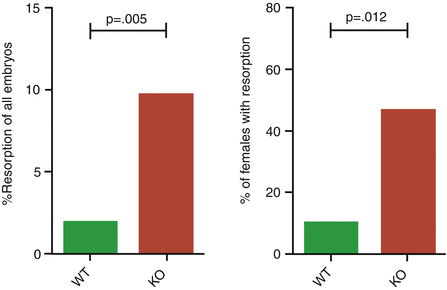
Fig. 4.2
Percent of resorbed embryos in CNS1-sufficient (wild type) and CNS1-deficient pregnancies in knockout mice
4.5 Dendritic Cells
Dendritic cells (DCs) are at the biological bridge between the innate and the adaptive immune system and their activation and modulation is critical for the outcome of the immune response. DCs localize in the peripheral tissue and act as sentinel of the immune response. After an inflammatory stimulus, DCs migrate via lymphatic vessels to the lymph node to present antigen to T cells [7]. According to their state (activated or not activated), DCs can secrete pro- or anti-inflammatory cytokines, thereby inducing immune responses or suppressing them, respectively.
In pregnancy, the number of DCs in the decidua is very low and the cells are phenotypically immature. The role of decidual DCs has not been fully elucidated; recently, it was suggested that DCs, as well NKs and macrophages, could act at the feto-maternal interface, intervening in the tissue remodeling and promoting maternal immune tolerance [7]. An increased number of phenotypically mature DCs have been associated with spontaneous miscarriage [24].
4.6 B Cells
B cells are a component of the humoral immunity of the acquired immune system. The main function of B cells is the production of antibodies (IgG, IgM, IgE, IgA), but B cells can also uptake, process, and present antigens and can produce several cytokines influencing immunity. The role of B cells during pregnancy has been poorly studied, but it was postulated that they may contribute toward pregnancy tolerance by regulating the production of the immune-modulatory cytokine IL-10 [25]. A physiological decrease of B cells was noted in the third trimester of normal pregnancy while preeclampsia has been associated with a persistence of a higher number of B cells. In women with autoimmune disease, it is possible that B cells compromise gestation, producing excessive autoantibodies [25].
4.7 Trophoblast as Regulator of Innate Immune Cell
Trophoblastic cells express pattern recognition receptors that act as “sensors” of the surrounding microenvironment [5]. These sensors allow the recognition of the presence of bacteria, viruses, damaged tissues, and dying cells by the trophoblast. Trophoblastic cells can secrete a specific set of cytokines influencing the immune cells within the decidua with the aim of instructing inflammatory cells to create an adequate microenvironment, to promote pregnancy evolution [5]. The success of the pregnancy depends on the trophoblastic ability to communicate with any immune cell type (monocytes/macrophages, pTreg and NK cells) and to coordinate their interrelated work [5, 26].
4.8 The Alteration of the Maternal–Fetal Equilibrium of the Inflammatory Cells
Alterations in type, number, aspect, or interrelationship of any component of the cells of the inflammatory milieu may be detrimental for pregnancy outcome. This might be the consequence of infections or of maternal immunological problems, either in clinical or in subclinical conditions.
Infection can reach the placenta and the fetus through the maternal blood circulation or by ascending into the uterus from the vagina or by descending into the uterus from the peritoneal cavity, by activation of the pathogenic phenotype of preexisting bacteria, or even by sudden immune reaction toward preexisting symbiotic bacteria.
4.8.1 Acute Chorioamnionitis
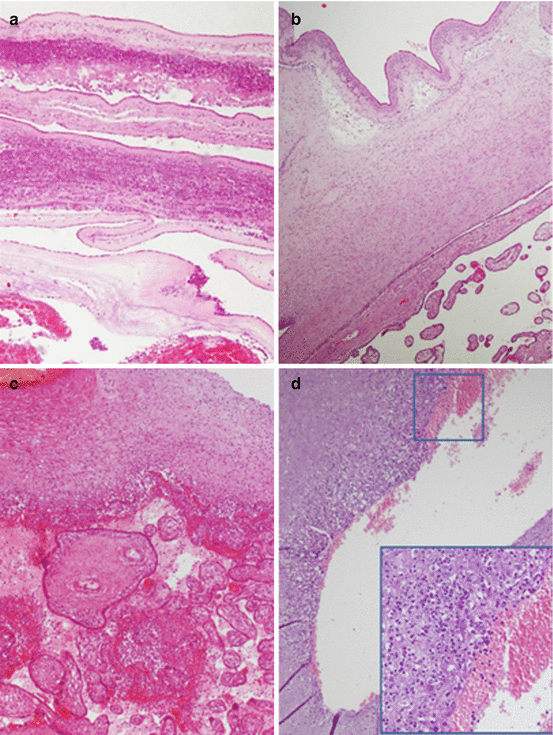
These processes could develop into acute chorioamnionitis. In these cases, most of infecting agents originate in the cervicovaginal tract and gain access to the uterine cavity breaching the normal cervical barrier. Maternal neutrophils infiltrate the chorionic plate and membranous chorioamnion (Fig. 4.3) [27]. Normally, neutrophils are a population of inflammatory cells that do not infiltrate placental compartments. When an infective insult occurs, neutrophils can invade placental structures; they can derive both from the intervillous space and the venules of decidua capsularis. In case of acute chorioamnionitis, a maternal and/or a fetal inflammatory response should be distinguished. The stage (progression of the disease) and the grade (intensity of the disease) of the inflammatory response are summarized in Table 4.2. Acute chorioamnionitis has been associated with adverse pregnancy and neonatal outcome, in relation to prematurity due to preterm delivery and/or preterm premature rupture of the membrane [29]. Fetal inflammatory response is the main factor that affected fetal–neonatal well-being; fetal inflammatory response involves chorionic plate vessels and/or umbilical vein prior to the involving of umbilical arteries. The association between fetal inflammatory response and adverse neonatal condition and outcome takes place especially when umbilical arteries are involved. Neonatal sepsis could be a consequence of fetal inflammatory response but also neurological impairment may develop including cerebral palsy [30].
 Nutrition Around the Clock and Inflammation: The Right Food at the Right Time Makes the Difference
Nutrition Around the Clock and Inflammation: The Right Food at the Right Time Makes the Difference
 Obesity and Inflammation in Pregnancy
Obesity and Inflammation in Pregnancy
 PCOS and Pregnancy: Impact of Endocrine and Metabolic Factors
PCOS and Pregnancy: Impact of Endocrine and Metabolic Factors
 The Role of Placental Inflammasomes in Linking the Adverse Effects of Maternal Obesity on Fetal Development
The Role of Placental Inflammasomes in Linking the Adverse Effects of Maternal Obesity on Fetal Development
 Gastrointestinal Symptoms and Nutritional Profile During Pregnancy
Gastrointestinal Symptoms and Nutritional Profile During Pregnancy
 Maternal Diet, Developmental Origins, and the Intergenerational Transmission of Cardiometabolic Traits: A Window of Opportunity for the Prevention of Metabolic Syndrome?
Maternal Diet, Developmental Origins, and the Intergenerational Transmission of Cardiometabolic Traits: A Window of Opportunity for the Prevention of Metabolic Syndrome?




Related posts:
Nutrition Around the Clock and Inflammation: The Right Food at the Right Time Makes the Difference
Obesity and Inflammation in Pregnancy
PCOS and Pregnancy: Impact of Endocrine and Metabolic Factors
The Role of Placental Inflammasomes in Linking the Adverse Effects of Maternal Obesity on Fetal Development
Gastrointestinal Symptoms and Nutritional Profile During Pregnancy
Maternal Diet, Developmental Origins, and the Intergenerational Transmission of Cardiometabolic Traits: A Window of Opportunity for the Prevention of Metabolic Syndrome?
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
