Chapter 7 Mononuclear Phagocytes in Immune Defense
• Macrophages: The ‘big eaters’. Macrophages are endowed with a remarkable capacity to internalize material through phagocytosis.
• Macrophages differentiate from circulating blood monocytes and are widely distributed throughout the body. Macrophages belong to the family of mononuclear phagocytes, which also comprise monocytes, osteoclasts, and dendritic cells. These cells share a common hematopoietic precursor that cannot differentiate into neutrophils. Phenotypically distinct populations of macrophages are present in each organ.
• Macrophages are highly effective endocytic and phagocytic cells. Macrophages have a highly developed endocytic compartment that mediates the uptake of a wide range of stimuli and targets them for degradation in lysosomes.
• Macrophages sample their environment through opsonic and non-opsonic receptors. Macrophages express a wide range of receptors that act as sensors of the physiological status of organs, including the presence of infection.
• Clearance of apoptotic cells by macrophages produces anti-inflammatory signals. Macrophages produce IL-10 and TGF-β upon internalization of apoptotic cells.
• Macrophages coordinate the inflammatory response. Recognition of necrotic cells and microbial compounds by macrophages initiates inflammation leading to the recruitment of neutrophils. Monocyte recruitment to sites of inflammation is promoted by activated neutrophils and there is a collaborative effort between macrophages and neutrophils to eliminate the triggering insult. Macrophages are actively involved in the resolution of the inflammatory reaction.
• There are different pathways of macrophage activation. TH1 cytokines such as IFNγ enhance inflammation and anti-microbial activity. TH2 cytokines induce an alternate activation that promotes tissue repair. TGFβ, corticosteroids and IL-10 can induce an anti-inflammatory phenotype.
Macrophages: the ‘big eaters’
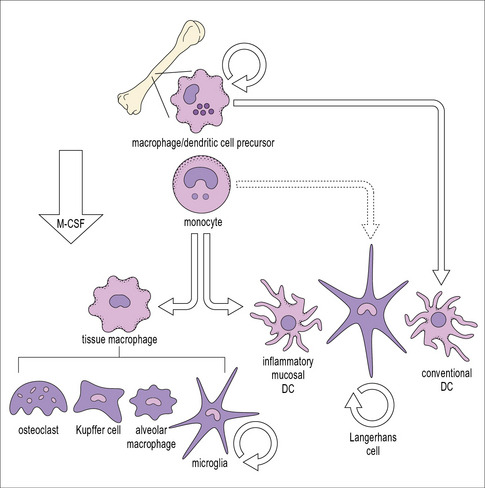
Macrophages differentiate from blood monocytes
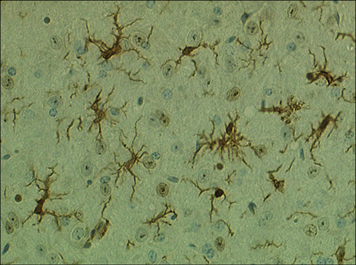
Within tissues the mononuclear phagocytes undergo maturation, adapt to their local microenvironment, and differentiate into various cell types (Fig. 7.1). Distinctive populations of resident macrophages are found in most tissues of the body; they differ in their life span, morphology, and phenotype, for example the microglial cells in the brain appear quite unlike mononuclear phagocytes in other tissues (Fig. 7.2). Resident cells have usually ceased to proliferate, but may remain as relatively long-lived cells, with low turnover, unlike neutrophils.

Macrophage populations have distinctive phenotypes
Some of the key features of resident macrophage populations in tissues are shown in Figure 7.w1. The study of the heterogeneity and distribution of mononuclear phagocytes has been made possible through the use of antibodies against differentiation markers. These markers can be located at the plasma membrane or endosomal compartments. In the mouse the F4/80 antigen and macrosialin (CD68) have proved useful in defining the distribution of mature macrophages in many (but not all) tissues. Differentiation antigens such as sialoadhesin, a lectin-like receptor for sialylated glycoconjugates, are particularly strongly present on populations of macrophages in lymphoid organs that do not express F4/80 or CD68. Macrophages expressing sialoadhesin are exposed to the blood and lymph and are thought to be involved in antigen delivery. In humans, the CD68 antigen, the human homolog of macrosialin, is widely expressed in macrophages while the F4/80 homolog EMR2 labels subsets of macrophages. Anatomical differences between mouse and human spleen correlate with different distribution of macrophage differentiation markers. The mannose receptor is expressed by macrophages in the red pulp of mouse spleen but is absent from these cells in human spleen.

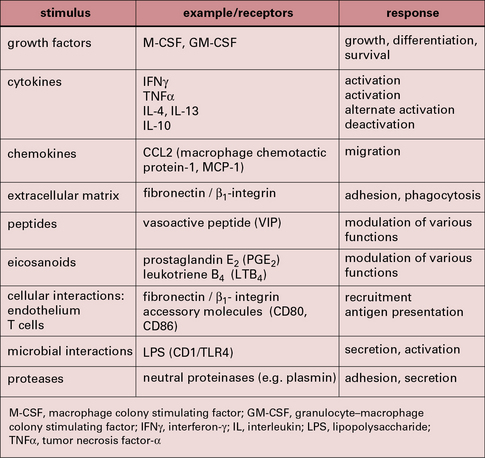
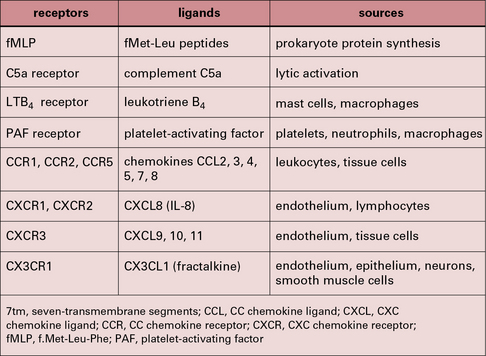
Some of the important stimuli that modulate macrophage phenotype are listed in Figure 7.w2 and some of their characteristic cell surface receptors in Figure 7.w3.
The tissue environment controls differentiation of resident macrophages
It is possible to reconstruct a constitutive migration pathway in which monocytes become endothelial-like and line vascular sinusoids, as in the liver (Kupffer cells, see Fig. 2.3), or penetrate between endothelial cells. They underlie endothelia or epithelia or enter the interstitial space or serosal cavities (Fig. 7.3). The molecular mechanisms of constitutive macrophage distribution and induced migration are beginning to be defined, and involve cellular adhesion molecules, cytokines, and growth factors, as well as chemokines and chemokine receptors.
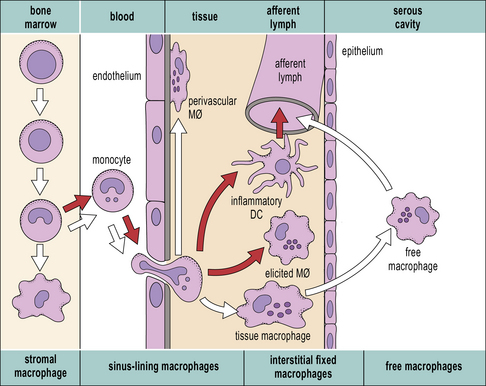
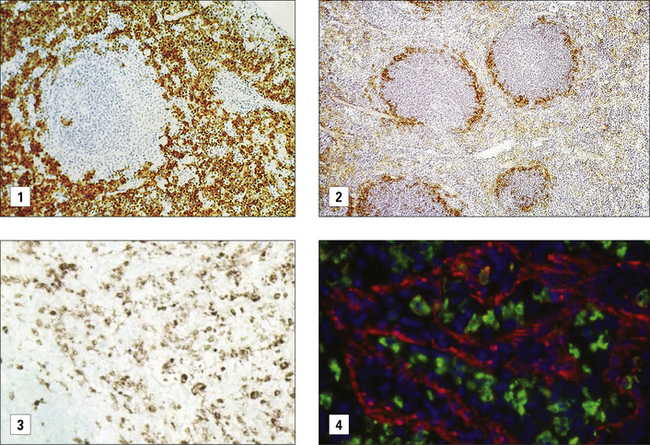
Secondary lymphoid organs contain several distinct types of macrophages. These macrophages have been better characterized in the mouse (Fig. 7.4) and subsets involved in the clearance of apoptotic lymphocytes (tingible body macrophages) or presentation of naive antigens to B cells (subcapsular sinus macrophages) have been identified. Anatomical differences between human and mouse spleen, such as the absence of a well defined marginal sinus, correlates with phenotypical differences in splenic macrophages (see Fig. 7.4)
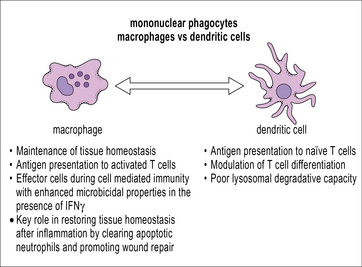
Macrophages can act as antigen-presenting cells
Macrophages, like dendritic cells, have all the machinery required for antigen processing and presentation of exogenous peptides and endogenous peptides on MHC class II and class I, respectively. Cross-presentation, a process by which peptides of exogenous origin are presented on MHC class I, also takes place in macrophages. While dendritic cells are uniquely suited for stimulating naive T cells in secondary lymphoid organs, macrophages present antigen in the periphery to activated (already primed) T cells. This interaction makes macrophages important effector cells during adaptive immunity (Fig. 7.5). The specialization of dendritic cells for antigen presentation correlates with a reduced degradative capacity that facilitates the generation of MHC-peptide complexes.
Macrophages act as sentinels within the tissues
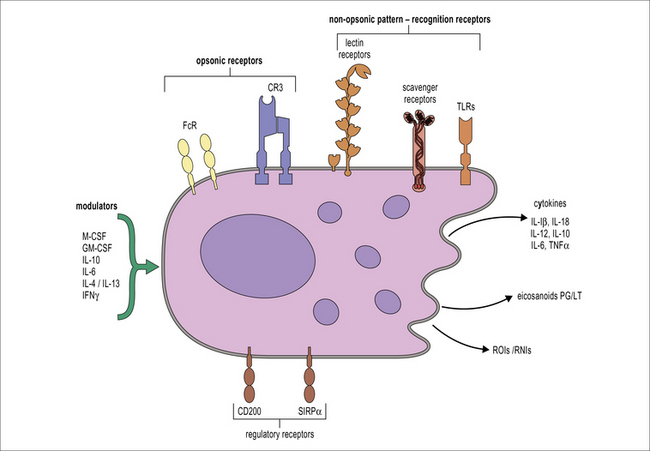
Macrophages react to a wide range of environmental influences that help to fulfill their role as sentinels of the innate immune system (Fig. 7.6). The presence of cells within tissues with the potential to initiate inflammation through the release of cytokines and chemokines and to cause tissue damage through the production of reactive oxygen species requires control systems capable of downmodulating macrophage activation. One of these systems involves the molecule CD200L, which is an inhibitory receptor expressed by myeloid cells. CD200L inhibitory signaling is triggered through interaction with CD200 expressed by non-hematopoietic cells as well as macrophages. The CD200–CD200L interaction is important for the control of macrophage activation by other cells present in tissues.
Phagocytosis and endocytosis
Large particles are internalized by phagocytosis
Phagocytosis involves the uptake of particulate material (>0.5 μm) after recognition by opsonic or non-opsonic receptors (see Fig 7.6), its engulfment through the generation of pseudopodia and the formation of phagosomes. Phagosomes follow a similar maturation process to endosomes through the fusion with components of the early and late endocytic compartments so that maturing phagosomes sequentially adopt characteristics of early and late endosomes; this process culminates in the fusion of phagosomes to lysosomes to form phagolysosomes (Fig. 7.7). Phagosomal maturation is accompanied by acidification of the lumen (from 6.1–6.5 in early phagosomes to 4.5 in phagolysosomes), which controls membrane traffic and has a direct effect on microbial growth. Other microbicidal mechanisms associated with phagosome maturation are the generation of reactive oxygen and nitrogen species and the presence of antimicrobial proteins and peptides.
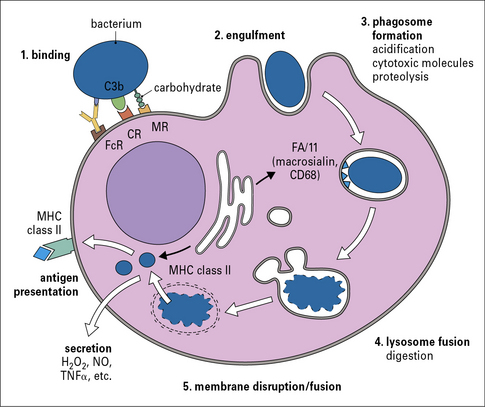
Macrophages sample their environment through opsonic and non-opsonic receptors
Opsonic receptors require antibody or complement to recognize the target
FcRs belong to the immunoglobulin superfamily (see Fig. 3.17). The best characterized FcR is CD64 (FcγRI), the high affinity receptor for IgG which signals through the common γ chain that contains an immunoreceptor tyrosine based activation motif (ITAM). The common γ chain is also used by some non-opsonic receptors that bind carbohydrates (see below) and it signals through the key kinase Syk. In humans other activating receptors for IgG are low affinity FcγRIIa (CD32) and FcγRIII (CD16), which require the recognition of immune complexes for inducing internalisation. IgG-opsonized material is readily internalized by macrophages and leads to the production of reactive oxygen species and cellular activation. The activating effect of ITAM-associated FcγRs is regulated by the presence of the inhibitory form of CD32 (FcγRIIb), which bears an ITIM.
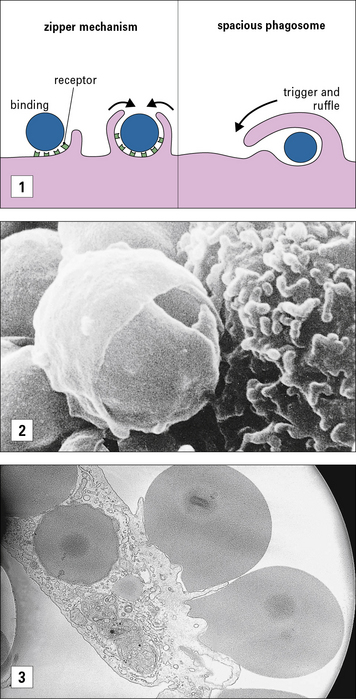
The mechanism for ingestion of antibody-coated particles is distinct from that mediated by CR3 (Fig. 7.8). FcR-mediated uptake proceeds by a zipper-like process where sequential attachment between receptors and ligands guides pseudopod flow around the circumference of the particle. In contrast, CR3 contact sites are discontinuous for complement-coated particles which ‘sink’ into the macrophage cytoplasm. Small GTPases play distinct roles in actin cytoskeleton engagement by each receptor-mediated process.
The best characterized non-opsonic receptors are the Toll-like receptors (TLRs)
Non-opsonic receptors or pattern recognition receptors (PRRs) recognize unusual features characteristic of damaged, malfunctioning or infected tissues and their general characteristics are described in Chapter 6.
TLRs are membrane glycoproteins with an extracellular region responsible for ligand binding and a cytoplasmic domain responsible for triggering an intracellular signaling cascade. They can form hetero- or homo-dimers with each other or complex with other receptors in order to detect a wide range of microbial components. They are located at the cell surface or within endosomes. In humans there are 10 of these receptors and together they are able to recognize a wide range of microbes including Gram-positive bacteria and mycobacteria (see Fig. 6.20). For example, TLR4 detects Gram-negative bacteria because of its ability to recognize endotoxin. It then signals to the cell using similar systems to those mediated by IL-1 (Fig. 7.9). It can also activate the macrophage by a second pathway that is initiated by Trif, which leads to a secondary production of IFNβ and autocrine activation of additional macrophage genes.
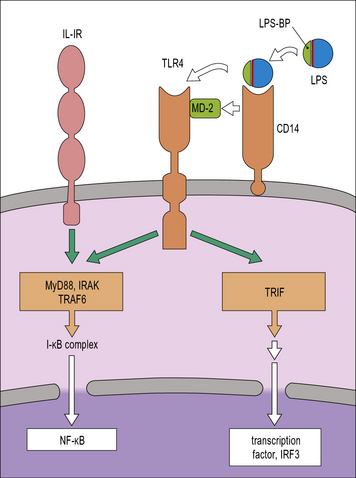
CD14 is a GPI-linked membrane protein that facilitates the recognition of LPS by TLR4 so that it increases LPS sensitivity (see Fig. 7.9). Recently CD14 has also been shown to facilitate recognition of ligands by TLR2 and TLR3, which opens the possibility of CD14 acting as a multifunctional adaptor protein.

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


