Chapter 1 Introduction to the Immune System
• The immune system has evolved to protect us from pathogens. Intracellular pathogens infect individual cells (e.g. viruses), whereas extracellular pathogens divide outside cells in blood, tissues or the body cavities (e.g. many bacteria and parasites). These two kinds of pathogens require fundamentally different immune responses.
• Phagocytes and lymphocytes are key mediators of immunity. Phagocytes internalize pathogens and degrade them. Lymphocytes (B and T cells) have receptors that recognize specific molecular components of pathogens and have specialized functions. B cells make antibodies (effective against extracellular pathogens), cytotoxic T lymphocytes (CTLs) kill virally infected cells, and helper T cells coordinate the immune response by direct cell–cell interactions and the release of cytokines.
• Specificity and memory are two essential features of adaptive immune responses. As a result, the adaptive arm of the immune system (B and T lymphocytes) mounts a more effective response on second and subsequent encounters with a particular antigen. Non-adaptive (innate) immune responses (mediated, for example, by complement, phagocytes, and natural killer cells) do not alter on repeated exposure to an infectious agent.
• Antigens are molecules that are recognized by receptors on lymphocytes. B cells usually recognize intact antigen molecules, whereas T cells recognize antigen fragments displayed on the surface of the body’s own cells.
• An immune response occurs in two phases – antigen recognition and antigen eradication. In the first phase clonal selection involves recognition of antigen by particular clones of lymphocytes, leading to clonal expansion of specific clones of T and B cells and differentiation to effector and memory cells. In the effector phase, these lymphocytes coordinate an immune response, which eliminates the source of the antigen.
• Vaccination depends on the specificity and memory of adaptive immunity. Vaccination is based on the key elements of adaptive immunity, namely specificity and memory. Memory cells allow the immune system to mount a much stronger and more rapid response on a second encounter with antigen.
• Inflammation is a response to tissue damage. It allows antibodies, complement system molecules, and leukocytes to enter the tissue at the site of infection, resulting in phagocytosis and destruction of the pathogens. Lymphocytes are also required to recognize and destroy infected cells in the tissues.
• The immune system may fail (immunopathology). This can be a result of immunodeficiency, hypersensitivity, or dysregulation leading to autoimmune diseases.
• Normal immune reactions can be inconvenient in modern medicine, for example blood transfusion reactions and graft rejection.
• illustrates how the components of the immune system fit together to allow students to grasp the ‘big picture’ before delving into the material in more depth in subsequent chapters;
• introduces the basic elements of the immune system and of immune responses, which are mediated principally by white blood cells or leukocytes (from the Greek for ‘white cell’) and are detailed in Chapters 2–12.
Cells and soluble mediators of the immune system
Cells of the immune system
Immune responses are mediated by a variety of cells and the soluble molecules that these cells secrete (Fig. 1.1). Although the leukocytes are central to all immune responses, other cells in the tissues also participate, by signaling to the lymphocytes and responding to the cytokines (soluble intercellular signaling molecules) released by T cells and macrophages.
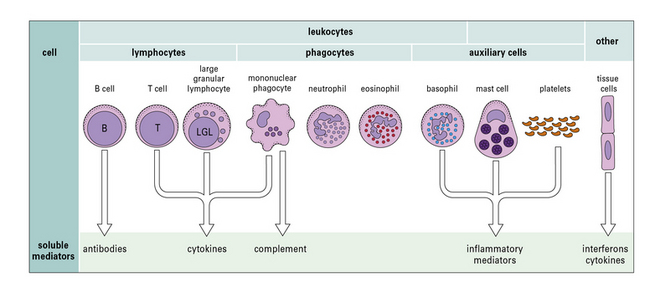
Fig. 1.1 Components of the immune system
The principal cells of the immune system and the mediators they produce are shown. Neutrophils, eosinophils, and basophils are collectively known as polymorphonuclear granulocytes (see Chapter 2). Cytotoxic cells include cytotoxic T lymphocytes (CTLs), natural killer (NK) cells (large granular lymphocytes [LGLs]), and eosinophils. Complement is made primarily by the liver, though there is some synthesis by mononuclear phagocytes. Note that each cell produces and secretes only a particular set of cytokines or inflammatory mediators.
Phagocytes internalize antigens and pathogens, and break them down
The most important long-lived phagocytic cells belong to the mononuclear phagocyte lineage. These cells are all derived from bone marrow stem cells, and their function is to engulf particles, including infectious agents, internalize them and destroy them. To do so, mononuclear phagocytes have surface receptors that allow them to recognize and bind to a wide variety of microbial macromolecules. They can then internalize and kill the micro-organism (Fig. 1.2). The process of phagocytosis describes the internalization (endocytosis) of large particles or microbes. The primitive responses of phagocytes are highly effective, and people with genetic defects in phagocytic cells often succumb to infections in infancy.
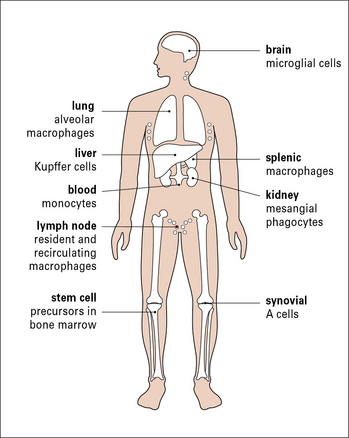
To intercept pathogens, mononuclear phagocytes are strategically placed where they will encounter them. For example, the Kupffer cells of the liver line the sinusoids along which blood flows, while the synovial A cells line the synovial cavity (Fig. 1.3).
B cells and T cells are responsible for the specific recognition of antigens
These two classes of lymphocytes carry out very different protective functions:
• B cells are responsible for the production of antibodies that act against extracellular pathogens
• T cells are mainly concerned with cellular immune responses to intracellular pathogens, such as viruses. They also regulate the responses of B cells and the overall immune response.
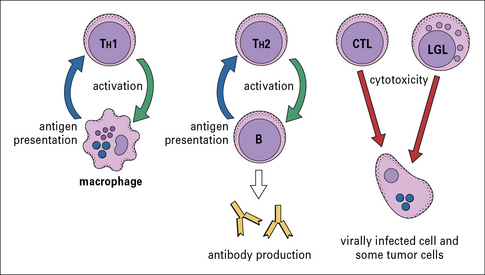
There are several different types of T cell, and they have a variety of functions (Fig 1.4):
• one group interacts with mononuclear phagocytes and helps them destroy intracellular pathogens – these are called type 1 helper T cells or TH1 cells;
• another group interacts with B cells and helps them to divide, differentiate, and make antibody – these are the type 2 helper T cells or TH2 cells;
• a third group of T cells is responsible for the destruction of host cells that have become infected by viruses or other intracellular pathogens – this kind of action is called cytotoxicity and these T cells are therefore called cytotoxic T lymphocytes (CTLs or TC cells).
Soluble mediators of immunity
Complement proteins mediate phagocytosis, control inflammation and interact with antibodies in immune defense
The complement system, a key component of innate immunity, is a group of about 20 serum proteins whose overall function is the control of inflammation (Fig. 1.5). The components interact with each other, and with other elements of the immune system. For example:
• a number of microorganisms spontaneously activate the complement system, via the so-called ‘alternative pathway’, which is an innate immune defense – this results in the microorganism being opsonized (i.e. coated by complement molecules, leading to its uptake by phagocytes);
• the complement system can also be activated by antibodies or by mannose binding lectin bound to the pathogen surface via the ‘classical pathway’.
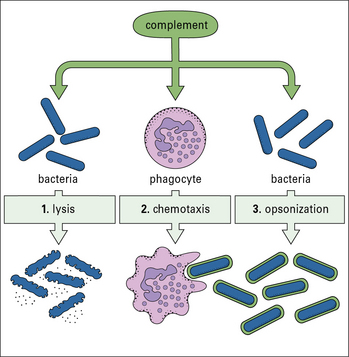
• opsonization of microorganisms for uptake by phagocytes and eventual intracellular killing;
• attraction of phagocytes to sites of infection (chemotaxis);
• increased blood flow to the site of activation and increased permeability of capillaries to plasma molecules;
• damage to plasma membranes on cells, Gram-negative bacteria, enveloped viruses, or other organisms that have caused complement activation. This can result in lysis of the cell or virus and so reduce the infection;
Cytokines signal between lymphocytes, phagocytes and other cells of the body
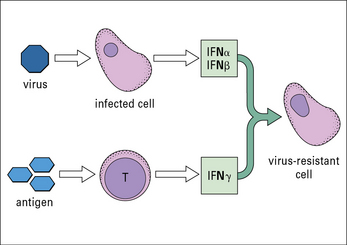
• one group of interferons (IFNα and IFNβ or type-1 interferons) is produced by cells that have become infected by a virus;
IFNs induce a state of antiviral resistance in uninfected cells (Fig. 1.6). They are produced very early in infection and are important in delaying the spread of a virus until the adaptive immune response has developed.