), Woodrow and Donohoe (1968); 2 ( ), E Borst-Eilers (personal communication); 3 (•), Poulain and Huchet (1971).
), E Borst-Eilers (personal communication); 3 (•), Poulain and Huchet (1971).
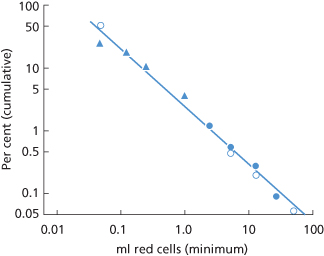
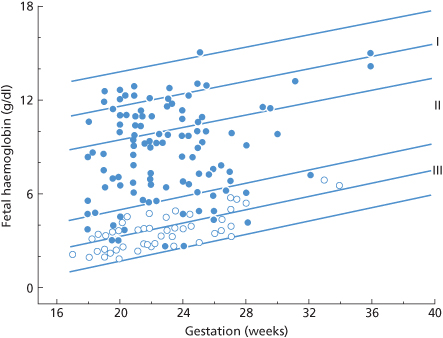
In Figure 12.1, the magnitude of TPH on a logarithmic scale is plotted against cumulative frequency, also on a logarithmic scale. The fact that the data can be reasonably well fitted by a straight line is convenient, as it is easy to read off the expected frequency of interpolated values. As the figure shows, about 1% of women have 3.0 ml or more of fetal red cells in their circulation at the time of delivery and 0.3% have 10 ml or more. The latter estimate agrees closely with other estimates, for example more than about 10 ml of fetal red cells in 0.2% of recently delivered women (Bartsch 1972) and 15 ml or more in 0.3%, based on about 8000 cases from seven different centres (Zipursky 1971).
The data plotted in Figure 12.1 suggest that the fact that fetal red cells can be detected in the circulation of only about 50% of recently delivered women, using the acid-elution method, is due simply to the relative insensitivity of the method.
Only women with ABO-compatible infants were included in the first series; in the other two series, the cases were unselected. Each point indicates the percentage of women found to have x ml or more of fetal red cells in their circulation immediately after delivery.
Effect of ABO Incompatibility
When the fetus is ABO incompatible with the mother, fetal red cells are less frequently detected in her circulation and then only in small numbers.
In two large series, when the fetus was ABO incompatible, fetal red cells were found in 19% and 24.7% of cases, respectively, compared with 50% and 56%, respectively, when the fetus was ABO compatible (Cohen and Zuelzer 1967; Woodrow and Donohoe 1968). Moreover, when the fetus was ABO incompatible, the amount of fetal red cells was usually very small, for example 0.1 ml or more in only 1.9% of cases when the fetus was ABO incompatible, compared with 18.5% when it was compatible (Woodrow and Donohoe 1968). In another series, 0.05 ml or more of red cells were found in 0.5% of cases when the fetus was ABO incompatible but in 23.1% of cases in which it was ABO compatible (Schneider 1969).
Caesarean Section and Manual Removal of Placenta
These are associated with a considerable increase in the number of fetal red cells found in the maternal circulation: in one series the incidence of those with fetal cells was 23.5% compared with only 5.2% in mothers who had had a normal labour (Finn et al. 1963), and in another series the number of women with more than 0.5 ml of fetal blood in the circulation was about 11% after a caesarean section or manual removal of the placenta compared with less than 1% in women with uncomplicated deliveries (Zipursky et al. 1963a,b). Similarly, of women with a TPH corresponding to 5 ml or more of fetal red cells, 12% had had the placenta removed manually compared with 1% of those who had not; fetal distress during labour was also significantly more frequent when there was a TPH of 5 ml or more (Li et al. 1988).
‘Late’ Entry of Fetal Red Cells into the Maternal Circulation, Via the Peritoneal Cavity
There seem to be two possible routes by which fetal red cells may reach the mother’s peritoneal cavity and thence make their way into her circulation. First, if they are spilled into the uterine cavity, they may pass into the peritoneal cavity by the fallopian tubes. Occasionally this can be observed after vaginal termination of pregnancy, when this is followed immediately by tubal ligation or diathermy. Second, fetal red cells may be spilt into the peritoneal cavity at hysterotomy or caesarean section. In one series of 46 women who underwent caesarean section, fetal cells were not detected in any maternal samples at 4 h but were found in six cases at 6 days (Hindemann 1966).
Massive Transplacental Haemorrhage
A massive TPH may be arbitrarily defined as one of more than 25 ml of red cells (50 ml of blood). TPHs of this magnitude occur in approximately one per 1000 pregnancies (Renaer et al. 1976) (see also Figure 12.1). If the blood volume of an infant at term is taken as 270 ml plus 120 ml in the placental circulation, i.e. 390 ml, a haemorrhage of 50 ml is equivalent to 13% of the circulating blood volume.
The cause of most massive TPHs is unknown, but can occasionally be identified, as when there is maternal trauma, for example a fall or a car accident, or when there is a placental chorioangioma (Sims et al. 1976) or a uterine choriocarcinoma (Blackburn 1976). From a review of the literature and from 64 personally observed cases, it was concluded that more than 50% of women with a TPH of 15 ml or more of fetal red cells have no history of any of the risk factors commonly believed to be important, for example intrapartum manipulation or placenta praevia. The few risk factors that were associated with large TPHs included premature separation of the placenta, manual removal of the placenta and fetal death (Sebring and Polesky 1990). Dziegiel and co-workers (2005) describe a case of massive TPH from an ABO-compatible fetus, which demonstrated the lifespan of some fetal red cells in the adult circulation to be the same as that of adult red cells.
When the fetus is ABO incompatible, a large TPH may be masked. In two cases in which infants were born with Hb concentrations of 3.5 and 6.9 g/dl, respectively, the percentage of fetal red cells demonstrable in the mother’s circulation was 0.6% and 1.9% respectively. As these infants must have lost amounts of red cells of the order of 100 ml, at least 6% of the cells in the mother’s circulation would have been expected to have been of fetal origin if they had survived normally, i.e. had been ABO compatible (Cohen and Zuelzer 1967).
Effect on Fetus
Of women who had had a stillbirth, 4.5% were found to have the equivalent of 25 ml or more of fetal red cells in their circulation (Huchet et al. 1975). This incidence is far higher than in a random series, leading to the conclusion that some stillbirths are caused by a massive TPH.
When a massive TPH occurs during delivery, the infant may be in a condition of oligaemic shock at birth but with a more or less normal Hb concentration, as there has been too little time for haemodilution. On the other hand, when there has been chronic severe bleeding the infant may be born with severe anaemia with signs of intense blood regeneration (Huchet et al. 1975; Renaer et al. 1976; Sims et al. 1976); very occasionally, hydrops has been observed (e.g. Weisert and Marstrander 1960). In one case, diagnosed from reduced fetal movements, haemorrhage was estimated as 350 ml of blood; the fetal PCV was 0.056. Following two intrauterine transfusions, the hydrops resolved and the infant was born alive. After further transfusions, the infant’s progress was normal (Konugres et al. 1994).
Chronic fetal–maternal bleeding may be responsible for iron deficiency anaemia in the fetus (Pearson and Diamond 1959; Miles et al. 1971).
Effect on the Mother
When the fetus is ABO incompatible, a large TPH may be associated with haemoglobinuria in the mother. One such episode followed an easy external cephalic version under general anaesthesia. The mother (group O) passed fetal Hb in the urine (0.2 g Hb/dl). Four weeks later, a group A infant was born; the placenta showed a small area of separation (Pollock 1968). In 14 further cases of external version, fetal cells were found in only one, and in this case the mother’s blood had not been examined before version.
The Passage of Red Cells from Mother to Fetus
The passage of red cells from mother to fetus is far less common than traffic in the reverse direction. Using a rosetting test, maternal red cells were detected in 70% of infants, but the total volume of maternal red cells in the infant exceeded 0.01 ml in only 2 out of 92 cases (Helderweirt 1963). With a new cytological method, the Kleihauer immunogold-silver-staining technique, 73 newborn infants were tested; the mothers had been selected as having an Rh antigen (D, c or E) absent from the infant’s red cells so that their cells could be recognized serologically; maternal red cells equivalent to 1 µl or more of blood were detected in only three infants; the amount was about 1 µl in two cases and about 100 µl in the third (Brossard et al. 1996).
Frequency of Opportunities for Rh D Immunization in Pregnancy
In about one of every 10 pregnancies in white people, the mother is D negative and the fetus is D positive. The calculation is as follows: 17% of women are D negative; 83% of their partners are D positive of whom 35 are DD and 48 Dd. All children born to the homozygous (DD) fathers will be D positive but only one-half of those born to the heterozygous (Dd) fathers will be D positive. Thus, of every 100 pregnancies, the number in which the fetus is D positive and the mother is D negative will be:
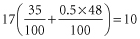
When primary Rh D immunization is induced in a first pregnancy, the amount of anti-D produced is almost always too small to produce haemolytic disease in the fetus. In considering the opportunities for producing Rh D haemolytic disease, one must therefore calculate how frequently a D-negative woman gives birth to two D-positive infants. As family size is often restricted to two children, one may consider how often a D-negative woman has two D-positive infants in succession. When her partner is DD, all her infants will be D positive, but when he is Dd, the chance that he will pass on D twice in succession is only one in four.
Thus, of every 100 second pregnancies, the number in which a D-negative woman is carrying her second D-positive infant is given by:
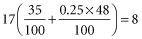
In calculating the frequency of opportunities for immunization to Rh D, account must be taken of the fact that primary immunization to Rh D is rare when the fetal red cells are ABO incompatible with the mother’s serum, as occurs, in white people, in about 20% of pregnancies. Thus, the figure ‘8’ must be multiplied by 0.8 (80%) to give an estimate of the number, i.e. about 6, of second pregnancies in every 100 in which a D-negative mother would be expected to have an infant with Rh D haemolytic disease if: (1) every first pregnancy with an ABO-compatible, D-positive infant induced primary immunization to Rh D; and (2) every subsequent pregnancy with a D-positive infant resulted in haemolytic disease. The frequency of Rh D haemolytic disease in the era before immunoprophylaxis was about one-sixth of this, i.e. 1 per 100 of second pregnancies. The main reasons why, in five out of six cases, in the absence of immunoprophylaxis, the second infant is not affected with haemolytic disease are, first, that the amount of TPH is often too small to induce primary immunization and, second, that even when the amount is large enough to induce primary immunization in some subjects, in others (non-responders) it is not.
In the preceding paragraph, for convenience, a positive DAT has been regarded as diagnostic of haemolytic disease. As discussed above, some infants with a positive DAT due to maternally derived alloantibody have no signs of increased red cell destruction. However, in practice there is no way of distinguishing in a newborn infant between no increased red cell destruction and a slight increase in red cell destruction and so, in calculating the frequency of the disease, all infants with a positive DAT have to be included.
Primary Rh D Immunization Caused by Pregnancy in the Absence of Immunosuppressive Therapy
Abortion
There is evidence that significant TPH occurs only after curettage and does not occur after either threatened or incomplete spontaneous abortions (Katz 1969; Matthews and Matthews 1969; Jørgensen 1975). In fact, evidence that spontaneous abortion occurring in the first trimester can cause primary Rh D immunization scarcely exists. In one series of women who had not been given anti-D Ig after an abortion occurring in the first 12 weeks of pregnancy and who developed anti-D during the following pregnancy, there were eight in whom the abortion had been spontaneous. In these eight cases, anti-D developed only during the last weeks of the succeeding pregnancy, and immmunization might well have been initiated by TPH occurring during the second pregnancy rather than by the previous abortion (LAD Tovey, personal communication).
Following induced abortion in D-negative primiparae, anti-D was found within the first 3 months of the following pregnancy in 1.5% and by term in 3.1% of the women (Simonovits et al. 1980). A similar frequency was found in another series, in which termination in the second month of pregnancy seemed to carry about the same risk of inducing immunization as termination carried out later (Hocevar and Glonar 1974). As 40% of fetuses carried by D-negative women are expected to be D positive, the frequency with which immunization to Rh D occurs following the induced abortion of a D-positive fetus must be higher than 3%. Even after making allowance for the fact that some women must become primarily immunized during the following pregnancy, the overall risk of immunization following the termination of pregnancy in a D-negative woman seems to be at least 4%.
Anti-D Present by the End of a First Full-Term Pregnancy
In D-negative women with no history of previous transfusion who are tested at the end of their first pregnancy with a D-positive infant, anti-D is found in approximately 1%, for example 0.8%, deduced from the data of Hartmann and Brendemoen (1953); 0.71%, (Eklund and Nevanlinna 1973); and 0.9%, i.e. 18 in 2000 (Tovey et al. 1983). An apparently much higher incidence, namely 62 in 3533, or 1.8%, was reported in a Canadian series (Bowman et al. 1978). However, the series included some women who had been pregnant previously and some in whom anti-D was not present at delivery although present 3 days later. If only primiparae are considered and only those in whom anti-D was found by the time of delivery, the figures become 34 in 2767, or 1.2% (excluding one woman in whom anti-D was detected at 11 weeks). The remaining discrepancy between this figure and those (0.7–0.9%) reported by others can be explained by the fact that, in almost one-third of the Canadian cases, the anti-D was detectable only in the autoanalyzer or by enzyme methods. If only cases in which the antibody was detectable by IAT are included, the figures become the same as those reported by others.
When anti-D develops during a first pregnancy, it is most commonly first detectable in the last few weeks: in one series, antibody was detected between 36 weeks and term in 50% (Eklund and Nevanlinna 1971); in another, antibody was detected in about 40% at 34 weeks, although not at 28 weeks, and in the remaining 60% was found for the first time at delivery, having been undetectable at 34 weeks (Tovey et al. 1983). In the Canadian series, among the 34 primiparae (see above), anti-D was first detected by 28 weeks in two, by 34 weeks in three more, between 35 and 40 weeks in 15 more, and immediately after delivery in the remaining 14; antibody was found in a further 10 for the first time 3 days after delivery (Bowman et al. 1978).
Cases in which anti-D was detected 8 days after delivery, having been undetectable at the time of delivery, were originally reported by Bishop and Krieger (1969) and were also noted by Jørgensen (1975).
The evidence that, in a substantial number of D-negative women, primary immunization to Rh D occurs during pregnancy indicates that anti-D Ig must be given antenatally as well as postnatally if Rh D haemolytic disease is to be prevented in as many cases as possible.
Anti-D First Detected 3–6 Months After a First Pregnancy, or During or After a Subsequent Pregnancy
Estimates of the incidence of anti-D in D-negative women 6 months after the birth of a first D-positive, ABO-compatible infant range from 4.3% in 1012 mothers (Eklund and Nevanlinna 1973) through 7.7% in 337 mothers (Borst-Eilers 1972) and 8.2% in about 400 mothers (Woodrow 1970) to 9.0% in 106 mothers (Jørgensen 1975). When antibody is present at 6 months it can almost always be detected at 3 months (Eklund and Nevanlinna 1973).
After the birth of a first D-positive infant to a D-negative mother, a relationship can be demonstrated between the number of fetal red cells demonstrable in the mother’s circulation at the time of delivery and the chance that anti-D will appear. When no fetal cells are detectable, anti-D is found in only about 3% of cases, whereas when the amount is 0.1 ml or more anti-D is found in about 31% of cases (Woodrow 1970). In the series of Woodrow (1970) in which 8.2% of D-negative mothers had anti-D in their plasma 6 months after their first pregnancy with a D-positive fetus, by the end of a second pregnancy with a D-positive fetus the frequency of anti-D was 17% (or one in six). The appearance of anti-D in a further 9% of women during the second pregnancy must have been due, in the great majority of cases, to a secondary response during the second pregnancy, so that the conclusion is that, for every woman who develops serologically detectable anti-D following a first pregnancy, there is another who has been primarily immunized but who requires a further stimulus to produce sufficient anti-D to be detectable serologically.
Anti-D developing during or following a second or later pregnancy may of course be due to primary Rh D immunization initiated during that pregnancy. Anti-D detected only during the last few weeks of pregnancy or after the pregnancy is likely to be due to primary immunization occurring during that pregnancy, whereas anti-D detected early in a pregnancy is likely to represent secondary Rh D immunization.
After delivery it is not uncommon for the titre of anti-D to increase, reaching a peak 1–3 weeks postpartum (Boorman et al. 1945).
Fetal Factors that May Affect Rh D Immunization
There is evidence that R2r infants are more effective in sensitizing their mothers to Rh D than are infants of other phenotypes (Murray 1957).
There is also evidence which suggests that the D-positive infant that initiates Rh D immunization is more frequently male than female. The sex ratio, M/F, in one series was 1.44 : 1, control 1.05 (Renkonen and Seppälä 1962), in another was 1.74 (Renkonen and Timonen 1967), and in another 1.5 (Woodrow 1970). In one more series, 18 out of 21 D-positive infants who initiated Rh D immunization in their mothers were male (Scott 1976).
Influence of ABO Incompatibility on Rh D Immunization in Pregnancy
The fact that immunization to Rh D during pregnancy is less common when the father is ABO incompatible with the mother was noted by Levine (1943). In a series of matings between D-positive fathers and D-negative mothers who had given birth to infants affected with HDN, 25% were ABO incompatible (father’s red cells vs. mother’s serum) compared with 35% of ABO-incompatible matings expected in the general population (for later reviews, see Nevanlinna 1965 and Levine 1958.) It has been estimated that (in white people) group A incompatibility between infant and mother gives 90% protection, and B incompatibility gives 55% protection against Rh D immunization (Murray et al. 1965).
Do Rh D-Negative Infants Get Immunized by Maternal Rh D-Positive Red Cells?
Although it has been claimed that some D-negative infants born to D-positive mothers develop anti-D within the first 6 months of life, the data have never seemed convincing (for references and discussion, see eighth edition, p. 653). Incidentally, tests on the maternal grandmothers of infants with HDN have failed to show any significant excess of D positives amongst them (Booth et al. 1953; Owen et al. 1954; Ward et al. 1957).
Clinical Manifestations of Rh D Haemolytic Disease
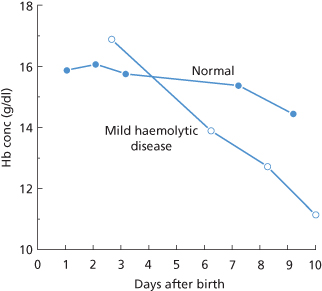
Haemolytic disease due to anti-D shows a wide spectrum of severity. In some D-positive infants whose red cells are coated with anti-D, as demonstrated by a positive DAT, there are no signs of red cell destruction. In infants in whom signs of red cell destruction are present, they may be so mild as to be recognized only by the development of mild jaundice on the first day or two of life and a slightly more rapid than normal fall in Hb concentration during the first 10 days or so (Figure 12.2). (In all newborn infants, the Hb concentration falls because red cell production decreases rapidly after birth, in association with a fall in erythropoietin levels, and remains depressed until the Hb concentration falls to about 10 g/dl.)
Figure 12.2 Rate of fall of haemoglobin (Hb) concentration in an infant mildly affected with haemolytic disease of the newborn, contrasted with the changes in a normal infant.
(Source: Mollison PL 1951. Reproduced with permission of John Wiley & Sons Ltd.) (from first edition, p. 385).
In more severely affected infants, jaundice may develop very rapidly (icterus gravis neonatorum). Unless the infant is treated promptly by exchange transfusion it may develop kernicterus, a syndrome characterized by lethargy, spasticity and opisthotonos, followed by irregular respiration. Of infants who develop kernicterus, about 70% die between about the second and fifth days of life and are found at autopsy to have yellow staining of the basal ganglia of the brain. Those who survive have permanent cerebral damage, characterized by choreoathetosis and spasticity; some infants show high-frequency deafness as the only sign. There is a close relationship between the peak serum bilirubin concentration and the development of kernicterus. In mature infants, kernicterus seldom, if ever, develops in association with serum bilirubin concentrations of less than 18 mg/dl (306 µmol/l) (Mollison and Cutbush 1951, 1954), although in premature infants there is some evidence that kernicterus develops at lower bilirubin levels (Ackerman et al. 1970).
In infants with a still more severe haemolytic process, profound anaemia develops and the infants may die in utero at any time from about the seventh week of gestation onwards.
In a series in which blood samples were obtained by fetoscopy at 18–24 weeks from 29 fetuses with severe haemolytic disease there were 14 with Hb values of less than 4 g/dl and 10 of these had the syndrome of hydrops fetalis (Nicolaides et al. 1985a). In this syndrome there is ascites and generalized oedema together with gross enlargement of the liver and spleen and, in many cases, of the heart. The oedema has been ascribed to the low serum albumin concentration that is almost always found (Phibbs et al. 1974). The finding of a relatively high concentration of albumin in the ascitic fluid suggests that damage to vascular endothelium, secondary to chronic hypoxia, may be an important feature in lowering the serum albumin concentration (Nicolaides et al. 1985b).
Severe hydrops fetalis is complicated by intravascular coagulation with widespread pulmonary haemorrhage and, in many cases, subarachnoid haemorrhage (Ellis et al. 1979). The outlook for infants with established or threatened hydrops due to haemolytic disease has been dramatically improved by the introduction of intravascular transfusion in utero (see below).
Non-immunological hydrops occurs in about 1 in 3500 births (MacAfee et al. 1970; Hutchinson et al. 1982). The list of causes is very long; among the commonest are cardiac abnormalities, chromosomal abnormalities and the twin-to-twin transfusion syndrome (Holzgreve et al. 1985). A single case has been described of hydrops due to a massive transfer, estimated at 470 ml of blood, from the maternal into the fetal circulation (Bowman et al. 1984).
The mortality rate of non-immunological hydrops is over 80% (Holzgreve et al. 1985). The subsequent infant is almost always normal (MacAfee et al. 1970).
Before the introduction of immunoprophylaxis with anti-D Ig, more than 80% of cases of fetal hydrops were due to haemolytic disease (MacAfee et al. 1970). Now, the percentage is less than 20.
Pattern of Haemolytic Disease in Successive Siblings
When anti-D develops during the first pregnancy it is only very seldom detected as early as the twenty-eighth week and is most commonly first detected during the last few weeks of pregnancy (see above). Moreover, when anti-D does develop during a first pregnancy, the concentration of the antibody is usually low. Accordingly, a D-positive infant born following a first pregnancy in a previous unimmunized woman very seldom shows clinical signs of haemolytic disease, and may simply have a positive DAT. When severe haemolytic disease is observed in a first-born infant it must always be suspected that the mother was immunized before the pregnancy.
In the first affected infant (i.e. in most cases born to a mother following a second or later pregnancy) haemolytic disease tends to be less severe than in subsequent affected siblings. For example, in first affected infants the stillbirth rate is 6% (Walker et al. 1957) but in second affected and later infants is about 29% (sixth edition, p. 677). After the second affected infant there is no tendency for the disease to become progressively more severe (HR Nevanlinna, personal communication, 1964; for further details, see previous editions of this book). Accordingly, in women who have had an affected infant the history is very helpful in giving a prognosis for any future infants. For example, the risk of stillbirth in a woman who has previously had a mildly affected infant is of the order of 2%, whereas in a woman who has had one previous stillbirth the risk of a subsequent stillbirth is as high as 70% (Walker et al. 1957). However, exceptions are known to occur. In one mother to whom a mildly affected infant was born after one fatally affected infant and two moderately affected ones, Fc receptor blocking antibodies were demonstrated in the mother’s serum (see below).
Significance of a Previous Transfusion to the Mother
Although the prognosis for infants of women who have previously been transfused with D-positive blood is not essentially different from that of other affected infants (Nevanlinna 1953), this generalization does not seem to apply if only first affected infants are considered (Levine et al. 1953; Mollison and Cutbush 1954). The greater severity of the disease in first affected infants born to previously transfused mothers is presumably due simply to the fact that a transfusion is a better primary stimulus than a small TPH.
Routine Tests to Detect Rh D Immunization
All so far unimmunized women should have their Rh D group determined on at least two occasions during pregnancy: at their first antenatal visit and at about 32 weeks.
Tests for Anti-D
The sera of all D-negative women should be tested for anti-D at the time of their first antenatal visit, i.e. at about the twelfth week of gestation. In women believed to be pregnant for the first time, anti-D will be found only occasionally, namely in those who have been immunized either by a previous transfusion of D-positive blood or by a previous unrecognized or undisclosed abortion. In women known to have been pregnant before, the presence of anti-D at 12 weeks will usually be due to the fact that they have not been given anti-D Ig following an earlier pregnancy or, if it has been given, the dose has been too small or given too late. In women without anti-D at their first antenatal visit, it is sufficient to make the next test at 28–32 weeks. As discussed later, there is a strong case for beginning immunoprophylaxis with anti-D Ig at 28 weeks. In women who have not formed anti-D when tested at 28–32 weeks, no further tests need be made until the time of delivery when, as soon as the infant is born, cord blood should be taken for determination of the D group and the DAT.
Changes in Anti-D Concentration When the Infant Is D Negative
Significant rises in anti-D concentration in women carrying a D-negative fetus were found in 4 out of 239 cases in one series (Hopkins 1970) and in 13 out of 300 in another (Fraser and Tovey 1976).
Antenatal Tests for Alloantibodies in D-Positive Women
When D-positive women form antibodies such as anti-c or anti-E, or alloantibodies outside the Rh system such as anti-K, there may be serious delay in diagnosing haemolytic disease in their infant after birth unless antenatal tests have revealed the presence of the antibody. The serum of all pregnant women should be screened for alloantibodies at the time of their first antenatal visit. If a clinically significant antibody is found, monthly tests until about 32 weeks are indicated, with two-weekly tests until term and a DAT on the infant’s cord blood at the time of delivery. If no alloantibodies are found at the first antenatal visit, it is sufficient to make a second test at 32 weeks.
Determination of D Group of Fetus in Utero
The D group of the fetus can be determined prenatally, by polymerase chain reaction (PCR) amplification of that region of fetal DNA encoding RHD. Initially, fetal DNA for this purpose was obtained from amniotic fibroblasts. Only 1–2 ml of amniotic fluid is needed and can be obtained late in the first trimester or early in the second. The test can also be done on a chorionic villus sample but this carries greater risks for the fetus and is also a cause of TPH (Bennett et al. 1993). The preferred method utilizes fetal DNA present in maternal plasma, as this poses no risk to the pregnancy or of TPH and thereby the possibility of sensitizing the mother to fetal antigens. Maternal plasma contains about 3% fetal DNA in the first trimester rising to 6% in the third trimester. The method can be undertaken from the beginning of the second trimester (Lo et al. 1998). Maternal plasma is isolated from blood collected into EDTA and then either centrifuged (16 000 g × 10 min, Chiu et al. 2001) or passed through a 0.2 micron filter to remove all cellular material within 48 h of collection to minimize the amount of maternal DNA in the sample.
Methods for the determination of the D group of a fetus require very careful design. RHD and RHCE are highly homologous genes and the D-negative phenotype can arise from a large number of different genetic mechanisms (see Chapter 5). Early methods of D typing from DNA were based on the assumption that RHD was absent from the DNA of D-negative individuals and primers for PCR were designed around regions of significant difference between RHD and RHCE, most notably in exon 7, the 3′ untranslated region of exon 10 and a 600-bp size difference in intron 4 (Arce et al. 1993; Bennett et al. 1993; Wolter et al. 1993). It soon became clear that although complete absence of RHD is by far the commonest cause of the D-negative phenotype in white people this is not true of other races (Chapter 5). A particular problem is detection of the so-called pseudo RHD frequently found in black people with D-negative red cells. About 67% of D-negative black people have the pseudo RHD. It can be detected by amplification of exon 4 which contains a 37-bp insert in the pseudo RHD and so can be diagnosed from the size of the PCR fragment obtained (Singleton et al. 2000). These authors describe a multiplex PCR based on exon 4 and exon 7 that can be used for routine diagnostic purposes (Figure 12.3). A further complication in D-negative black Africans is that 15% have a hybrid gene RHD-CE-Ds which contains exons 1, 2, some or all of 3, 8, 9, 10 of RHD but exons 4, 5, 6, 7 from RHCE (Daniels et al. 1998). However this allele is easily detected by absence of exon 4 of RHD. Others have devised multiplex assays in which all RHD-specific coding exons are amplified in a single tube (Maaskant-van Wijk et al. 1998) and in which all RHD exons are amplified specifically and sequenced (Legler et al. 2001). The last two methods require optimal standardized conditions and are not so easily introduced into routine diagnostic laboratories (Van der Schoot et al. 2003).
Figure 12.3 Fetal D typing by PCR.
(Source: Singleton et al. 2000. Reproduced with permission of the American Society of Hematology.)
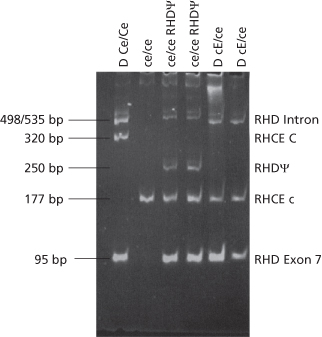
The D group of a fetus can also be determined using real-time PCR and this is the method preferred by most laboratories. This method requires two sequence-specific oligonucleotide primers to amplify the region of interest and a third oligonucleotide (probe) that anneals to a region within the amplicon. The probe has fluorescent dyes attached at its 5′-end (reporter dye) and its 3′-end (quencher dye). When the probe is intact, the proximity of the two dyes causes the emission spectrum of the reporter dye to be quenched. As the PCR progresses, the 5′ exonuclease activity of the Taq DNA polymerase causes cleavage of the reporter dye from the probe and the reporter dye fluorescence is detected by a laser. As the release of reporter dye fluorescence is directly proportional to the target amplification the method is quantitative (Figure 12.4). The method is extremely sensitive and, provided the PCR is designed to take account of common forms of D-negative phenotype where RHD is not completely absent such as the pseudo RHD and RHD-CE-Ds discussed above, can be used to provide a very effective diagnostic service (Finning et al. 2002; Daniels et al. 2009). One problem with this method concerns the availability of controls to ensure fetal DNA is present in cases where the D type is D negative. This is easily solved when the fetus is male by amplifying the Y chromosome-specific gene SRY. An international reference reagent is available for this purpose (Metcalfe et al. 2012). However, suitable markers when the fetus is female are not available. One approach is to test maternal DNA (derived from Buffy coat cells) for a number of insertion/deletion polymorphisms and then look for the presence of polymorphisms in fetal DNA that are absent from the mother’s DNA (Finning et al. 2004). Another method exploits differences in the degree of methylation of CpG islands in the 5’ of the tumour suppressor gene RASSF1A. Maternal DNA but not fetal DNA is digested with the methylation-sensitive restriction enzyme BstU1 (Chan et al. 2006). Quantification of total plasma DNA can be obtained by amplification of a housekeeping gene like CCR5 and reference to a calibration curve. If total DNA is greater than 125 ng/ml, white cell lysis may have occurred and the assay may be compromised by excessive maternal DNA. Quantification of total plasma DNA is also valuable for avoiding the problem of a D-negative mother with pseudo RHD, as quantification of the maternal gene will give values approaching the total DNA value, far higher than that found for fetal DNA sequences (Finning et al. 2004).
Mass spectrometry can be used to detect PCR products derived from fetal RHD with an accuracy comparable to that obtained using real-time quantitative PCR (Grill et al. 2009). A meta-analysis of 37 publications reporting results of fetal DNA testing utilizing DNA from maternal blood concluded the overall accuracy of this method is 94.8% (Geifman-Holtzman et al. (2006). However, Daniels et al. (2009) consider laboratories providing routine testing services achieve much greater accuracy and quote their own experience of testing 1400 pregnancies which yielded only two false-negative and four false-positive results. A false-positive result may be obtained if the mother has received a solid organ transplant. Minon et al. (2006) describe a case in which a mother had received a kidney transplant from a D-positive male and gave birth to a D-negative female. DNA isolated from maternal plasma predicted a D-positive fetus whereas DNA from amniotic fluid predicted a D-negative fetus.
Determination of RHD Zygosity
Knowledge of the genotype of the father can be helpful when anti-D is found in maternal serum or when a previous pregnancy has resulted in HDN. Serological methods are of limited value in this respect but several molecular methods have been described (reviewed in Van der Schoot et al. 2003). Real-time PCR can be used to compare the amount of RHD product with that of a reference gene (Krog et al. 2007). Another approach targets the so-called Rhesus box sequences upstream and downstream of RHD and RHCE respectively (see Figure 5.2). When RHD is deleted by an unequal crossover event, the crossover point occurs within the Rhesus boxes creating a hybrid Rhesus box comprising parts of the upstream and downstream boxes (Wagner and Flegel 2000). Assays that target the hybrid Rhesus box can therefore be used to determine zygosity (Chiu et al. 2001; Van der Schoot et al. 2003). The assays described above are not suitable for measurement of zygosity in non-white individuals
Antenatal Assessment of Severity
The severity of haemolytic disease can be assessed most reliably by fetal blood sampling but the procedure carries a small risk to the fetus and may provoke a TPH, followed by an increase in maternal antibody concentration. Accordingly, various non-invasive methods of assessment are used in the first instance and fetal blood sampling is used only in selected cases.
Estimation of Maternal Antibody Concentration
Estimations of antibody concentration, using an autoanalyzer, are useful in two ways. First, a low value almost always indicates that the infant will be mildly affected, or unaffected. In one series, in 78 cases in which the maternal anti-D concentration remained below 4 iu/ml (0.8 µg/ml), no infant had a cord Hb concentration of less than 100 g/l and only three infants needed an exchange transfusion (Bowell et al. 1982). Unfortunately, though, antibody levels of 6 iu/ml or more, predicting moderately severe disease, are quite commonly found when the infant is unaffected (Hadley et al. 1993).
Second, an increase in anti-D concentration is a warning that the severity of the haemolytic process may be increasing. In 11 cases in which fetal blood sampling had shown that the fetus was not anaemic, and in which maternal anti-D concentration rose to more than 15 iu/ml within the following 2–3 weeks, a further fetal sample showed that the fetus had become moderately or severely anaemic (Nicolaides and Rodeck 1992). In interpreting apparent changes in antibody concentration, it is essential to compare the latest sample with earlier samples by testing the samples together.
Relation between IgG Subclass of Anti-D and Severity
When only IgG3 anti-D was present, severity was less than when IgG1 alone or IgG1 + IgG3 were present (Zupanska et al. 1989; Pollock and Bowman 1990; Alie-Daran et al. 1992). In one investigation, when both IgG1 and IgG3 were present, IgG1 was usually preponderant and most severe disease correlated with IgG1, particularly G1m(3) (Alie-Daran et al. 1992). In other investigations, when only IgG1 was detected disease was more severe (Parinaud et al. 1985), less severe (Zupanska et al. 1989) or no different (Pollock and Bowman 1990) than when both IgG1 and IgG3 were present. These apparent discrepancies are probably due to the fact that when both IgG1 and IgG3 are present, the fraction of IgG1 may vary from 11% to 99% (Garner et al. 1995). Correlation of the amount of IgG1 and the severity of HDN is substantially better than that of IgG3 and severity (Garner et al. 1995; Lambin et al. 2002).
Cellular Bioassays in the Assessment of Severity
Bioassays are substantially more cumbersome to perform than estimates of antibody concentration but are expected on theoretical grounds to give better predictions of severity. Monocyte monolayer assays (MMAs), applied to maternal serum, have been found to distinguish between mildly and severely affected infants (Nance et al. 1989a; Zupanska et al. 1989). MMAs were more reliable than amniotic fluid measurements in predicting the need for treatment (Nance et al. 1989b).
In a series in which the results of an antibodydependent, cell-mediated, monocyte-dependent cytotoxicity [ADCC(M)] assay were compared with IAT titres, either at 32 weeks or at term, severe disease was correctly predicted in 75% by ADCC(M) but only in 15% by IAT titre (Engelfriet and Ouwehand 1990). In a small series in which an ADCC assay with lymphocytes (ADCC(L) assay) was used, of 10 women, all of whom had anti-D concentrations of 20 iu/ml or more, three had low assay values and these were the only ones whose infants did not require exchange transfusion (Urbaniak et al. 1984).
In a study in which samples were tested ‘blind’ by several different laboratories, the frequency of results with various assays that correctly predicted the severity of HDN was as follows: ADCC(M), 60%; ADCC(L), 57%; chemiluminescence (CL), 50%; adherence and phagocytosis with peripheral blood monocytes, 41% and with U937 cells or cultured macrophages, 32%. In many cases, maternal samples were taken only some time after delivery, which may explain why the frequencies of correct results were not higher (Report from Nine Collaborating Laboratories 1991).
In perhaps the most decisive investigation so far, sera from 44 D-negative women with D-positive infants were tested extensively; the series included approximately equal numbers of unaffected, moderately affected, severely affected and very severely affected infants. The tests used were the monocyte monolayer assay (MMA), antibody-dependent cellular cytotoxicity (ADCC) assay (with monocytes), estimation of anti-D concentration in the autoanalyzer and measurement of the number of IgG1 and IgG3 molecules bound to target cells. ADCC correctly predicted severity in 39 out of 44 cases, autoanalyzer quantification in 35 cases and MMA in 32 cases. ADCC activity and HDN severity were correlated with the number of IgG1 molecules bound to target cells but MMA activity was most closely correlated with the number of IgG3 molecules bound, thus explaining the superior predictive value of ADCC compared with MMA (Garner et al. 1995).
Unfortunately, cellular bioassays are cumbersome and are unsuitable for routine use.
False-Positive Results in Cellular Bioassays
In three circumstances, the fetus may be unaffected or only mildly affected despite a strongly positive result in a cellular bioassay:
Amniocentesis in Estimating the Severity of Haemolytic Disease
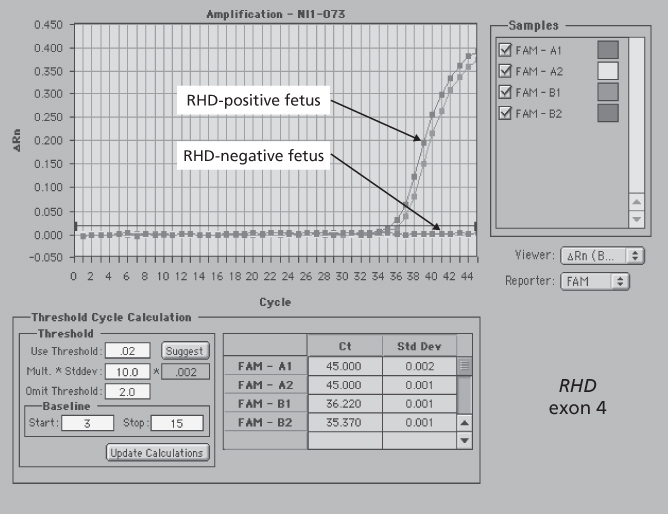
Estimation of the amount of bile pigment in amniotic fluid is performed by measuring the difference in optical density at 450 nm (OD450) between the observed density and an extrapolated baseline (Figure 12.5). The optical density of amniotic fluid falls during the last 13 weeks of pregnancy so that the stage of pregnancy has to be taken into account when interpreting the findings. Figure 12.6 shows a chart produced by Liley (1961) to indicate the approximate severity of disease for a particular OD450 at any stage of pregnancy from 27 weeks onwards.
Figure 12.5 Plot of optical density readings of amniotic fluid from an Rh D-immunized woman at the thirty-fifth week of pregnancy. The optical density at 450 nm (OD450) is expressed as the height (in this case 0.206) above the conjectural baseline.
(Source: Adapted from Bowman JM 1965. Reproduced with permission of the American Academy of Pediatrics.).
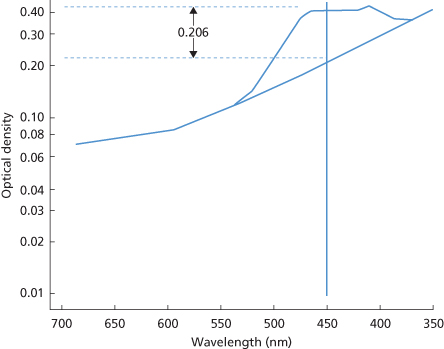
Figure 12.6 Liley’s three zones, indicating the approximate severity of haemolytic disease, from readings of the OD450 of amniotic fluid (see Figure 12.3) during the last trimester of pregnancy. Zone 1 indicates a mildly affected or unaffected infant, whereas zone 3 indicates a high probability of hydrops and fetal death; the significance of zone 2 is intermediate. In the original paper the chart begins, as here, at 27 weeks 1 day, i.e. at the start of the twenty-eighth week of gestation. Subsequently, the lines were extrapolated to 20 weeks but then provided a far less reliable guide (see text).
(Source: Liley 1961. Reproduced with permission of Elsevier.)
If the lines in Liley’s chart are extrapolated to earlier stages of pregnancy, misleading results are obtained. For example, in one series, if results had been interpreted in this way, 10 out of 31 severely anaemic fetuses (Hb <6 g/dl) would not have received an intrauterine transfusion, whereas many mildly affected fetuses would have been treated unnecessarily (Nicolaides et al. 1986).
Examination of the Fetus by Ultrasonography and Fetal Doppler Blood Flow Velocity
Ultrasonography can be used to diagnose hydrops fetalis as indicated by ascites, pleural or pericardial effusions and skin oedema (Bowman 1983). In fetuses of 18–24 weeks, ascites diagnosed in this way is always associated with Hb levels of less than 4 g/dl (Nicolaides et al. 1985a). Repeated examination of the fetus by ultrasonography can detect early signs of cardiac decompensation, i.e. a small pericardial effusion or dilatation of cardiac chambers.
Combined with monitoring of the fetal heart rate, sonographic examination makes conservative management of the fetus possible and helps to avoid unnecessary intervention (Frigoletto et al. 1986). In a study of 50 Rh-immunized pregnancies at 18–20 weeks’ gestation, when ultrasound was compared with fetal blood sampling, ultrasound was found unreliable in predicting fetal anaemia in the absence of hydrops (Nicolaides et al. 1988a).
Many studies report the value of Doppler velocimetry measurement of fetal middle cerebral artery peak systolic velocity in detecting moderate to severe fetal anaemia in red blood cell alloimmunized pregnancies. Mari and co-workers (2000) report an increase in peak systolic velocity to have a predictive value of 100% in moderate to severe anaemia in the presence or absence of hydrops. Others report similar findings (Dukler et al. 2003; Pereira et al. 2003). A comparison of Doppler velocimetry with determination of OD450 in amniotic fluid concluded the methods have similar test accuracy for predicting fetal anaemia and since Doppler velocimetry is non-invasive, this method carries less risk (Bullock et al. 2005). However, a systematic review of non-invasive techniques (fetal ultrasonography and Doppler blood flow velocity) to predict fetal anaemia, published in 2001, concluded ‘the literature reporting non-invasive techniques to predict fetal anaemia is methodologically poor and a standard approach to the evaluation of these techniques is lacking. A recommendation for practice cannot be generated without further rigorous research’ (Divakaran et al. 2001). Abdel-Fattah and Soothill (2002, cited in Petz and Garratty (2004), p. 524) conclude that careful attention to obstetric history and serial antibody quantitation is equally as important as Doppler velocimetry.
Fetal Blood Sampling
The preferred method of obtaining blood from the fetus in utero is by inserting a needle percutaneously and guiding it by ultrasonographic monitoring into the umbilical or hepatic vein. This procedure is described as cordocentesis or percutaneous umbilical vein sampling (PUBS). In 394 women in whom 606 fetal samplings were carried out by this method before the twenty-fourth week, the pregnancy loss rate was 0.8% (Daffos et al. 1985).
It is essential to confirm that the sample obtained is of pure fetal blood. A rapid method is to determine the mean cell volume (MCV) in an automated blood counter; fetal values are 118–135 µ3. Examination of a blood film stained by the Kleihauer–Betke method (see Appendix 13) is a more accurate method of excluding the presence of maternal red cells but takes substantially longer. Another reliable method is to visualize the streaming of microbubbles in the umbilical vein during the infusion of saline (King and Sacher 1989).
It is now routine practice to determine the fetal PCV or Hb concentration before intrauterine transfusion. Figure 12.7 shows the Hb concentration from 17 weeks to term of 106 non-hydropic and 48 hydropic infants with Rh D haemolytic disease. The figure also shows the range of values in 200 normal fetuses and 10 cord samples obtained at the time of delivery. As the figure shows, the Hb concentration of normal fetuses was found to increase from 11 g/dl at 17 weeks’ gestation to about 13.5 g/dl at 32 weeks and to 15.0 g/dl at term, although this latter value is somewhat lower than the generally accepted value of about 16.5 g/dl for the cord blood of normal mature infants.
Figure 12.7 Haemoglobin concentrations of 48 hydropic (s) and 106 non-hydropic (d) fetuses with HDN due to anti-D, at the time of taking the first sample from each fetus. Zone I indicates the zone of normal values (the lines mark the mean and limits of ± 2SD), based on samples from 200 normal fetuses undergoing prenatal tests. Zone III indicates the mean ± 2SD for hydropic fetuses, and zone II is that of moderate anaemia.
(Source: Nicolaides et al. 1988a. Reproduced with permission of Elsevier.)
A paper giving more details of the 110 fetuses in this series sampled between 15 and 21 weeks gave the following values before 22 weeks: for Hb, between 12.3 and 13.0 Hb g/dl, and for PCV, 0.373–0.393; there was no obvious trend towards higher values during this period although values for six fetuses at 15 weeks were slightly lower (Millar et al. 1985). In another series, of 163 fetuses of 18–30 weeks’ gestation, Hb increased from 11.47 g/dl at 18–20 weeks to 13.35 g/dl at 26–30 weeks; corresponding figures for PCV were 0.359–0.415 (Forestier et al. 1986).
The severity of haemolytic disease in utero can be assessed from the deviation of the observed Hb from the normal mean for the period of gestation. Normal fetal Hb can be predicted from: [85 + (1.4 × gestational weeks)] g/l, based on cordocentesis results in 726 pregnancies in which the fetus proved to be normal (Nicolaides and Rodeck 1992); a deficit of 2 g/dl is diagnosed as mild, of 2–7 g/dl as moderate, and of >7 g/dl as severe (Nicolaides et al. 1988b).
Routine Antenatal Determination of Fetal D Type
The availability of molecular methods for the determination of fetal D type using cell-free fetal DNA in maternal blood (see above) makes it feasible to consider the use of automated DNA typing of antenatal patients so that only those carrying a D-positive fetus are given prophylactic anti-D. Two studies report the use of high throughput fetal DNA typing using robotic isolation of DNA and real time quantitative PCR. In a study from the Netherlands the results of fetal RHD determination using an exon 7 probe at 30 weeks gestation on 1257 individuals gave three false-negative and five false-positive results (van der Schoot et al. 2006). A study from the UK used probes for RHD exon 5 and exon 7 and CCR5 as an amplification control. In this study of 1869 pregnancies the correct fetal D phenotype was predicted in 95.7% of cases (Finning et al. 2008). The first national ante-natal screening programme was implemented in Denmark (Clausen et al. 2012). Maternal blood samples from 2312 pregnancies were assayed in gestational week 25. The accuracy of testing was 96.5%. Unnecessary use of prophylactic anti-D was avoided in 862 cases.
Assessment of Severity in the Newborn Infant
Although many infants born with a positive DAT require no treatment, others need exchange transfusion or phototherapy if they are to be prevented from developing kernicterus. It is therefore essential to assess severity at the earliest possible moment. As explained below, there is a special advantage in testing cord blood rather than blood obtained from the infant after birth.
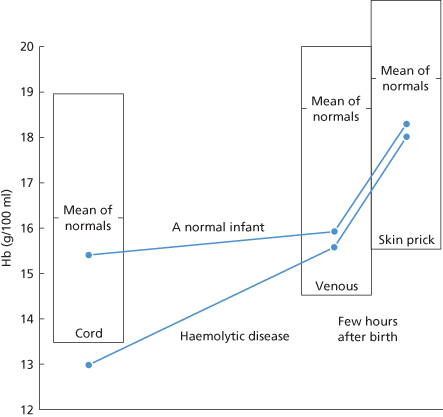
The best single criterion of severity of haemolytic disease is the cord Hb concentration, although in practice other criteria are always taken into account. The reason for preferring to test cord blood to blood obtained from the infant after birth is simply that during the few minutes after delivery a variable amount of blood is transferred from the placenta to the infant (Budin 1875; DeMarsh et al. 1942; Yao et al. 1969). Accordingly, interpreting Hb values in infants shortly after birth presents just the same difficulties as interpreting the significance of Hb values in any other subject who has recently been transfused with an unknown amount of red cells. The postnatal rise in Hb concentration has the effect of widening the normal range of values. Whereas the normal range (mean ± 2 SD) of Hb values in cord blood is approximately 13.6–19.6 g/dl, the range in samples taken on the first day of life is approximately 14.5–22.5 g/dl (Mollison and Cutbush 1949a). These figures refer to venous samples. In newborn infants, skin-prick samples tend to give distinctly higher values (for references, see third edition, pp. 579–80). The practical importance of Hb changes in the immediate postnatal period is illustrated in Figure 12.8. In the case shown, an infant with moderately severe haemolytic disease had a cord Hb concentration of 12.8 g/dl, a value definitely below the lower limit of normal, whereas a sample taken a few hours after birth was within the normal range.
Figure 12.8 Changes in haemoglobin (Hb) concentration produced by the placental transfer of blood tend to mask mild degrees of anaemia in newborn infants. Often the Hb concentration of skin-prick samples is much higher than that of venous samples and this is a second factor that may mask anaemia. In the case illustrated here, an infant with haemolytic disease of the newborn has a cord Hb concentration below the normal range, but after birth its Hb concentration is as high as that of many normal infants.
(Source: Mollison and Cutbush 1949a. Reproduced with permission of the BMJ Publishing Group.)
Before the introduction of immunoprophylaxis, almost 50% of affected infants had cord Hb concentrations of 14.5 g/dl or more, 30% had cord Hb values between 10.5 and 14.4 g/dl, and about 20% had Hb values of between 3.4 and 10.4 g/dl (second edition, p. 464). The probability of survival diminishes as the cord Hb concentration falls (Mollison and Cutbush 1951; Armitage and Mollison 1953); there is also a correlation between cord bilirubin concentration and severity, although the relation is less close than that between cord Hb concentration and severity (Mollison and Cutbush 1949a).
Indications for exchange transfusion and phototherapy are described below.
Estimation of the Amount of Antibody on Red Cells of Affected Infants
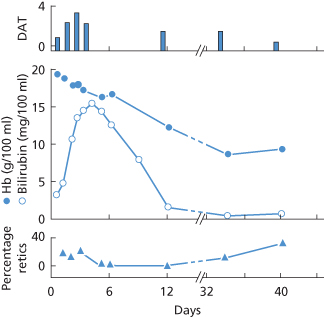
In one series, the amount of anti-D on the cord red cells varied from 0.4 to 18.0 µg/ml of cells. The amount of antibody on the red cells was not highly correlated with either the infant’s cord Hb concentration or the cord serum bilirubin concentration (correlation coefficient about 0.6). All 13 infants with more than 8 µg of anti-D per millilitre of red cells required treatment, but even at a level of 2 µg of anti-D per millilitre, 6 out of 14 infants required treatment (Hughes-Jones et al. 1967). In cases in which, due to a misunderstanding, a dose of 150–300 µg of anti-D has been given to a D-positive infant, instead of to its D-negative mother, only a very mild haemolytic syndrome has as a rule been produced. In one such case the infant’s red cells were shown to be coated with approximately 1.5 µg of anti-D per millilitre (Marsh et al. 1970; Figure 12.9).
Figure 12.9 Changes in haemoglobin (Hb) concentration (•—•); serum bilirubin concentration ( —
— ); reticulocyte count (
); reticulocyte count ( —
— ); and direct antiglobulin test (DAT; positive reactions graded from 0 to 4) in a D-positive infant injected, 5 h after birth, with 200 µg of anti-D.
); and direct antiglobulin test (DAT; positive reactions graded from 0 to 4) in a D-positive infant injected, 5 h after birth, with 200 µg of anti-D.
(Source: Marsh et al. 1970. Reproduced with permission of John Wiley & Sons Ltd.)
Antenatal Treatment of Haemolytic Disease
Plasma Exchange in the Mother
Although at first sight intensive plasma exchange in women immunized to Rh D seems entirely rational and might be expected to lead to a substantial lowering of the concentration of anti-D, to the benefit of the D-positive fetus in utero, results of this treatment have in fact been very variable. In one series of Rh-immunized women in whom a mean volume of 3.5 1 of plasma was exchanged on average twice a week, the anti-D level was rapidly lowered and maintained at a lower level than previously in 15 out of 35 cases. In the remaining 20, the anti-D level remained high despite intensive plasma exchange or was lower only until the thirtieth week of pregnancy, after which an uncontrollable rise occurred (Robinson 1984). Animal experiments indicate that one reason for lack of success is the fact that temporary lowering of antibody levels tends to provoke a rebound, suggesting that plasma antibody levels play an important part in regulating antibody production (Bystryn et al. 1970). The ability to maintain normal concentrations of serum IgG is critically dependent on FcRn, which maintains a balance between recycling of IgG and catabolism of IgG (Roopenian et al. 2003). It seems likely that removal of IgG by plasma exchange is compensated by an increase in IgG recycling through FcRn and a consequent reduction in the rate of catabolism.
In centres with a large experience of intravascular transfusion of the fetus in utero, plasma exchange has a very limited application because deaths in utero from haemolytic disease are believed not to occur before about 17 weeks, and i.v. transfusion becomes possible as early as the eighteenth week of gestation (Nicolaides and Rodeck 1985). It has been suggested that plasma exchange should be reserved for the woman with a previous history of hydrops developing before 24–26 weeks’ gestation, with a homozygous (DD) partner. Intensive plasma exchange should then be begun at 10–12 weeks, with fetal blood sampling at 19–22 weeks (Bowman 1990).
Absorption of Alloantibodies on to Red Cells
As mentioned in Chapter 1, one method of removing alloantibodies is to withdraw plasma, absorb the plasma with appropriate red cells and return it to the donor. In a case in which this method was used in an Rh-immunized woman there was a dramatic increase in anti-D concentration, which may well have been due to the presence of red cells or stroma in the plasma (Robinson 1981). To prevent this hazard, plasma can be passed through a microfilter (Yoshida et al. 1981).
Intravenous Immunoglobulin Given to the Mother
A single course totalling 2 g/kg over 5 days or repeated weekly injections of 1 g/kg have been tried in conjunction with plasma exchange or with intravascular transfusion of the fetus in two very small series. There was apparent benefit in one but not in the other (Rubo and Wahn 1990; Chitkara et al. 1990). In two much larger series, there was suggestive evidence that the severity of haemolytic disease in the fetus was reduced (Margulies and Voto 1991; C De la Camera and R Arrieta, personal communication). However, because there was great variability in previous obstetric histories, in the stage of pregnancy at which treatment was started and in the number of courses of Ig given, definite conclusions cannot yet be reached. In a more recent study, seven alloimmunized pregnant patients with a history of perinatal loss and two with markedly elevated maternal antibody titres were given serial plasmapheresis followed by weekly infusions of intravenous Ig, resulting in reduced maternal red cell antibody titres. Serial peak middle cerebral artery velocities remained below the threshold for moderate to severe fetal anemia during therapy and all seven infants survived (Ruma et al. 2007).
IVIG might act by saturating FcRn and thereby inhibiting placental transfer of anti-D to the fetus: in perfusion experiments with a lobule of human placenta, a plasma level of 20 g/l IgG in the maternal circulation, produced by injecting commercial IVIG, inhibited the transfer of the alloantibody anti-PlA1 (HPA-1a) (Morgan et al. 1991). Inhibition of placental anti-D IgG transfer was also observed at similar levels of maternal IgG (Urbaniak et al. 1999).
IVIG Given to the Fetus
In a small randomized trial, severely affected fetuses were treated with intravascular transfusions with, or without, IVIG in a mean dose of 85.7 mg/kg/body weight determined by ultrasound. IVIG conferred no apparent benefit, but the largest dose that could be given was too small and the average number of treatments was only two or three (Dooren et al. 1994).
Transfusion of the Fetus in Utero
Intraperitoneal transfusion was the first method to be used (Liley 1963): red cells are injected into the peritoneal cavity of the fetus, whence they are taken up into the bloodstream via the subdiaphragmatic lacunae and right lymphatic duct. Uptake is dependent on diaphragmatic movements (Menticoglou et al. 1987) and intraperitoneal transfusions are therefore useless in those hydropic fetuses who have no such movements. Although intraperitoneal transfusion has been largely superseded by intravascular transfusion, it still has a place. It is easy to perform and may also be used in combination with intravascular transfusion to increase the total volume of blood given to the fetus, thus prolonging the interval between transfusions (Nicolini and Rodeck 1988; Rodeck and Letsky 1989).
Intravascular transfusion is performed through a needle inserted into the umbilical vein by ultrasonographic guidance. Results obtained with this method have been spectacular. In one early series of 29 fetuses severely affected with Rh D haemolytic disease (10 with hydrops) treated at 18–24 weeks, 25 survived (Rodeck et al. 1984). In another series, 16 out of 22 hydropic fetuses survived and three of the six deaths were at 19–22 weeks in the fetuses of i.v. drug abusers (Bowman 1990). In a third series, the survival rate of fetuses developing hydrops before 26 weeks was 61%, and of those developing hydrops later was 100% (Grannum and Copel 1988).
In 10 published series of fetuses with severe haemolytic disease treated by intravascular transfusion in utero, of those with hydrops 66 out of 96 (69%) survived, compared with 114 out of 134 (85%) of those without hydrops. Hydrops was reversed in 60% of cases and in these, the survival rate was 92%, compared with 43% in those cases in which hydrops was not reversed (Tannirandorn and Rodeck 1990). Even in very experienced hands, a single intravascular transfusion has a mortality rate of 2% (Rodeck and Letsky 1989). In a retrospective study of pregnancy outcome following intrauterine transfusion for fetal anaemia due to red cell alloimmunization in the Netherlands during the period 1988–99, 210 fetuses from 208 pregnancies received 593 transfusions. Survival rate in fetuses without hydrops was 92% compared with 78% in those with hydrops. In maternal D immunizations, 89% of fetuses survived. The overall fetal loss rate was 4.8% (Van Kamp et al. 2004).
It is customary to use blood that is less than 72 h old. The red cells should be group O, D negative, K negative and should be compatible with the mother’s serum; the PCV of the red cell preparation should be 0.70–0.85. To avoid the risk of graft-versus-host disease, the red cells should be irradiated. The white cells should be removed by filtration to avoid possible immunomodulatory effects of transfusion and the transmission of cell-associated viruses such as cytomegalovirus (CMV) and human T-lymphotropic virus (HTLV). In addition to being submitted to all the mandatory tests, the blood should be screened for HBs and anti-CMV. The usual transfusion volume is 50 ml per kg estimated non-hydropic weight, reduced to 30–40 ml/kg estimated non-hydropic weight for hydropic fetuses (Bowman 1994). Red cells suspended in non-protein solutions such as SAG-M or Adsol should not be used for transfusion to the fetus in utero.
Alloimmunization of the mother may follow intrauterine sampling or transfusion of the fetus. Fetal or donor red cells may enter the maternal circulation, either by being shed into the mother’s peritoneal cavity or by entering maternal blood vessels in the placenta, and may stimulate the formation of ‘new’ alloantibodies or increase the concentration of existing ones.
In two series of Rh D-immunized women managed differently, when fetal blood sampling was used, other alloantibodies were found during pregnancy in 3 out of 63 cases and in a further 2 out of 38 examined after delivery; in contrast, when only amniotic fluid examinations were used, other antibodies were found during pregnancy in 1out of 52 cases and in 1 out of 22 examined after delivery (Pratt et al. 1989). The development of alloantibodies within 2–4 weeks of giving intraperitoneal transfusions to fetuses has been recorded in three cases: anti-Fyb in a woman who had already formed anti-Ce (Contreras et al. 1983), and anti-Jka and a combination of anti-Fyb, -Jkb and -S in two other women (Barrie and Quinn 1985).
Following a series of 68 fetal intravascular transfusions, TPH, as judged by an increase of at least 50% in the level of maternal α-FP, occurred in 27 cases (40%). The frequency was much higher (66%) in women with an anterior placenta than in those in whom the placenta was posterior or fundal (frequency of TPH, 17%). The mean estimated volume of TPH was 2.4 ml (blood). When the volume of TPH following the first intravascular transfusion exceeded 1 ml, the mother’s anti-D titre rose by more than 150% within the following 3 weeks (Nicolini and Rodeck 1988).
Donor red cells are less frequently the source of immunization than fetal red cells. Among 91 women, following 280 intrauterine transfusions (IUTs), additional antibodies appeared in 24 (26%). The immunizing source could be identified in 14; in 11 of these, the antibodies were directed against fetal antigens and in three, against donor antigens (Viëtor et al. 1994). Similar findings are reported by Schonewille et al. (2007) who studied 212 pregnancies in which donor red cells used for intrauterine transfusion were matched for Rh and Kell antigens. Twenty-five percent of women formed new antibodies and 48% of the new antibodies were directed at Rh and Kell antigens so presumably induced by the fetus. Evidence of induction of antibodies against antigens present on donor cells but not on the fetus was obtained in 11 cases, prompting the authors to consider the potential value of more comprehensive blood group phenotyping of donor red cells for intrauterine transfusions. High throughput DNA-based blood group genotyping of large donor inventories would facilitate this (see Chapter 8).
Induction of GvHD
In occasional infants transfused in utero, a subsequent postnatal exchange transfusion induces GvHD. The cells causing the GvHD have the HLA group of the donor used in the exchange transfusion and it is presumed that the IUT has an immunomodulatory effect (Williamson and Warwick 1995).
Premature Delivery
At a time when premature delivery was being practised only occasionally, it was found that approximately 50% of all stillbirths due to haemolytic disease occurred after 35 weeks’ gestation (Allen 1957; Walker et al. 1957). Because of the high mortality rate in premature infants at that time, there was reluctance to carry out premature delivery before about the thirty-fifth week of pregnancy but, with the steadily increasing success in the care of premature infants, delivery is now carried out as early as 30–32 weeks with high survival rates.
Postnatal Treatment of Haemolytic Disease
Exchange Transfusion
Exchange transfusion with D-negative blood greatly increases the survival rate and almost removes the risk of kernicterus (Allen et al. 1950; Mollison and Walker 1952). In the method introduced by Diamond (1947), blood is withdrawn and injected, intermittently, through a plastic catheter passed up the umbilical vein. The primary object of exchange transfusion is to remove D-positive red cells which, in a severe case of HDN, may have a survival time as short as 2 or 3 days (Mollison 1943). (In contrast, D-negative red cells almost always survive normally (Mollison 1951, p. 398).) The secondary object of exchange transfusion is to remove bilirubin already present in the plasma.
It is possible to predict accurately the volume of D-positive red cells remaining in the infant’s circulation if the infant’s initial PCV and body weight are known: a nomogram was published by Veall and Mollison (1950). However, the amount of bilirubin removed is more difficult to calculate, as during exchange transfusion bilirubin enters the plasma from the extravascular space. For this reason most clinicians like to exchange relatively large volumes of blood, for example 200 ml/kg, and to spend some time, for example 60–90 min, over the exchange transfusion.
Plasma-reduced blood with a PCV of at least 0.60 should be used for exchange transfusion; the blood should be group O, D negative and K negative, and should be compatible with the mother’s serum; it should be less than 7 days old. The blood should be screened for HbS and for anti-CMV, as well as being submitted to all the usual tests.
Erythropoiesis may be suppressed for many months after exchange transfusion and infants may be misdiagnosed as having red cell aplasia.
Indications for Exchange Transfusion
When the fetus has been transfused prenatally for severe disease, exchange transfusion is not usually required after birth and phototherapy (see below) is, in 80% of cases, adequate to control hyperbilirubinaemia. Exchange transfusion is carried out only if the serum bilirubin concentration threatens to reach or exceed 340 µmol/l.
Infants who have been transfused in utero and who do not require exchange transfusion after birth invariably require one or more simple ‘booster’ transfusions during the first 6–8 weeks of life. Hb concentration should be estimated at 10- to 14-day intervals until it stops falling. If the concentration falls towards 70 g/l, a transfusion of 20 ml of packed red cells/kg body weight should be given (JM Bowman, personal communication). In newborn infants who have not been transfused prenatally, a cord Hb concentration of 110 g/l is an indication for exchange transfusion. Before the introduction of improved methods of phototherapy, a cord serum bilirubin concentration of 60–70 µmol/l (in an infant with a cord Hb of 110 g/l or less) was also an indication for exchange transfusion but, now, phototherapy is tried first (JM Bowman, personal communication, 1996).
Phototherapy to Reduce Serum Bilirubin Concentration
On exposure to light, particularly in the region of 420–480 nm, bilirubin is converted to the non-toxic pigment, biliverdin. Exposing jaundiced newborn infants to a suitable fluorescent light lowers serum bilirubin concentration (Cremer et al. 1958; Costa et al. 1960), an effect that was confirmed in a controlled trial (Tabb et al. 1972).
Phototherapy, when applied early enough and with sufficient intensity, can avoid the need for exchange transfusion in many infants. One recent improvement in the efficacy of phototherapy has been the introduction of a fibreoptic blanket that delivers phototherapy to the anterior and posterior surfaces of the infant simultaneously. With this device the hourly decline in serum bilirubin concentration in low-birthweight infants with hyperbilirubinaemia was almost doubled (Holtrop et al. 1991).
Intravenous IgG to Reduce the Need for Exchange Transfusion
In a trial, infants with Rh HDN were randomly assigned as soon after birth as possible to receive phototherapy alone or phototherapy plus high-dose IgG (500 mg/kg over a 2-h period). Serum bilirubin was monitored 6-hourly. Exchange transfusion was required in 11 out of 16 infants receiving only phototherapy but in only 2 out of 16 who also received IgG (Rubo and Wahn 1991). A systematic review of randomized and quasi-randomized controlled trials comparing the effectiveness of high-dose intravenous immunoglobulin and phototherapy with phototherapy alone in neonates with Rh and/or ABO incompatibility concluded that significantly fewer infants required exchange transfusion in the intravenous immunoglobulin group (Gottstein and Cooke 2003).
Suppression of Rh D Immunization that Would Otherwise Follow Pregnancy
As pointed out in Chapter 5, Rh D immunization can be prevented by giving 20 µg or more of anti-D immunoglobulin for every 1 ml of D-positive red cells introduced into the circulation. So far as Rh D immunization by pregnancy is concerned there are two causes of failure to prevent immunization when anti-D immunoglobulin is given: either the dose of anti-D is insufficient or it is given too late, that is to say after primary Rh D immunization has been induced.
When anti-D immunoglobulin was first used to prevent Rh D immunization associated with pregnancy, it was injected immediately following delivery. It was assumed that if given during pregnancy it would harm the fetus and that, in any case, TPH occurred mainly during delivery so that treatment immediately after delivery would be effective. It eventually became clear that in a minority of cases primary Rh D immunization occurs during pregnancy so that when anti-D is injected only postnatally there is a regular, although small, failure rate.
The results of giving anti-D only postnatally will be considered first.
Administration of Anti-D Only Postnatally
Failure Rates at 6 Months
When 100–300 µg of anti-D Ig is injected immediately after delivery, the number of D-negative women who develop anti-D within the following 6 months is 0.1–0.5%. For example, 34 out of 33 260 (0.1%) in Finland, where women were treated with 250 µg of anti-D (Eklund, 1978); 55 out of 16 142 (0.34%) women treated in Scotland with either 100 or 200 µg of anti-D (I Cook, personal communication), and 16 out of 3113 (0.51%) in an Australian series. The somewhat higher failure rate in the Australian series does not seem to have been due to the inclusion of women already immunized before being treated, as anti-D was detected in 0.71% at delivery and these women were excluded (Davey 1976a).
Failure Rates at the End of a Second D-Positive Pregnancy
From what has been said already, the minimum failure rate expected is about 1.5%, i.e. an incidence of 0.7% in each of the two pregnancies due to primary immunization occurring during the pregnancy and an additional 0.2% or so for failures due to a TPH at the time of the first delivery too large to be covered by a dose of 300 µg of anti-D or less. In fact, failure rates of the order expected have been observed in many series; for example, 1.86% (see Davey (1976a), supplemented by personal communication]; approximately 1.5% (Eklund 1978); and approximately 1.5% in women treated with 50, 100 or 200 µg of anti-D in a controlled trial (see Table 12.1).
Table 12.1 Controlled trials of anti-D dosage in suppressing Rh D immunization (MRC 1974).
| Dose* (µg) | % of women with anti-D† | |
| 6 months after first pregnancy | At the end of second pregnancy | |
| 200 | 0.2 | 1.5 |
| 100 | 0.2 | 1.1 |
| 50 | 0.4 | 1.5 |
| 20 | 1.4 | 2.9 |
All women D negative with two D-positive infants, the first being ABO compatible.
* Anti-D injected i.m. within 36 h of delivery of first infant.
† Detected by indirect antiglobulin test.
When anti-D is detected for the first time during a second pregnancy in a woman who has been given anti-D Ig after her first pregnancy, there are two possible explanations: (1) sensitization to D occurred by the time of the first delivery but anti-D was produced in detectable amounts only after the further stimulus of a second pregnancy; (2) primary immunization to D occurred only during the second pregnancy. It seems that the second explanation is usually the right one; the evidence is that in these cases the antibody develops only towards the end of the second pregnancy, for example in the last 4 weeks in 50% of the cases in one series (Eklund 1978).
Effect of Different Amounts of Anti-D Given Immediately After Delivery
In a ‘blind’ trial conducted by a working party of the Medical Research Council (MRC) and begun in 1967 in the UK, different doses of anti-D were given to four groups of women (about 450 women in each group). Over the dose range 200 to 20 µg there was a significant, although small, trend towards an increase in the failure rate 6 months after a first pregnancy (see Table 12.1). Approximately 200 women in each of the four groups were followed through a second pregnancy with a D-positive infant; the differences in the failure rates in the different groups were not statistically significant, although failures were suggestively more frequent with the 20-µg dose than with the larger doses.
Doses of Anti-D for Postpartum Injection
Evidence that 20 µg of anti-D/ml red cells is effective in suppressing primary immunization has been reviewed earlier. To allow a margin of safety, 25 µg/ml has been recommended (WHO 1971). A dose of 300 µg of anti-D has long been the standard postpartum dose in the USA and many other countries, although 100 µg has been used in the UK. A dose of 300 µg covers TPHs of about 15 ml or less, that is to say the amounts found in approximately 99.8% of women at the time of delivery, whereas 100 µg covers TPHs of 4 ml or less, as found in about 99% of women. The dose of 300 µg was chosen to cover the vast majority of TPHs and to make screening tests for the occasional very large TPH inessential; nevertheless, a screening test capable of testing TPHs of 15 ml or more of red cells is mandatory in the USA (AABB 1991). With the 100-µg dose, the screening test is obviously even more important and must detect TPHs of 4 ml or more. Furthermore, when screening tests are positive, an estimate must be made of the size of the TPH so that an appropriate extra dose of anti-D can be given.
Flow cytometry can be used both to detect TPHs of 2 ml or more (see above) and to provide an accurate estimate of their size, but this technique is not widely available. In detecting TPHs of 15 ml or more, the rosetting test is probably the most convenient and can be used to provide semi-quantitative estimates. The acid-elution technique can also be used to provide quantitative estimates (Appendix 13).
In women in whom an unusually large TPH has been detected and who have been given extra anti-D Ig, tests are sometimes done to confirm that an adequate dose has been given. One practice is to look for the presence of anti-D in the maternal plasma but this is unsound in principle, as even when the antigen concentration is low, not all the antibody will be bound, however little is given. Furthermore, the method has been shown to be of no practical value (Ness and Salamon 1986). The method that seems most likely to be worthwhile is to test for clearance of fetal red cells from the maternal circulation, for example by using a rosetting test. Although a relationship between clearance and immunosuppression has yet to be firmly established, the two seem at least to be associated; see Chapter 3 for further discussion.
Doses of Anti-D Following Premature Termination of Pregnancy
In women undergoing termination of pregnancy, usually at about 3 months, a common practice is to give 50 µg of anti-D. In a series of 3080 women treated with this dose following therapeutic abortion, only 13 (0.42%) were found to be immunized during a second pregnancy (I Simonovits, personal communication), suggesting a very low failure rate of suppression.
The dose which should be given between about 12 and 20 weeks is uncertain; some give the same amount as for a termination at 12 weeks and others give the same dose as would be given at 28 weeks. There is general agreement that from 20 weeks onwards the dose used should be that normally used at 28 weeks (e.g. in the USA, 300 µg).
Antenatal Administration of Anti-D
As described above, a single pregnancy with a D positive, ABO-compatible infant initiates primary Rh D immunization in about one in six (17%) of D-negative women. When anti-D immunoglobulin is given postnatally, the incidence of primary Rh D immunization falls to about 1.5%, as judged by the development of anti-D by the time of delivery of a second D-positive infant. Thus, in some 90% of cases Rh D immunization that would otherwise follow pregnancy can be prevented by giving anti-D postnatally. As described below, it seems likely that most of the remaining 10% of cases can be prevented by antenatal treatment.
Safety of Antenatal Treatment
Although it was at first believed that injecting anti-D into a D-negative woman pregnant with a D-positive fetus was potentially dangerous, further consideration and practical experience show that the fear is groundless.
In D-positive infants born to mothers who are actively immunized to Rh D, the total amount of anti-D in the infant is only about 10% of that in the mother (Hughes-Jones et al. 1971). Accordingly, when 300 µg of anti-D is injected into the mother, even if equilibration across the placenta occurred immediately, not more than about 30 µg would be expected to be in the infant. In fact, as discussed earlier, IgG is transferred relatively slowly across the placenta. Accordingly, by the time that equilibrium is reached a considerable amount of the anti-D which has been injected into the mother will have been catabolized and the total amount reaching the infant will be less than 30 µg.
In trials in which 300 µg of anti-D were given to D-negative women at the twenty-eighth week and again at the thirty-fourth week of pregnancy, although as many as 28% of ABO-compatible D-positive infants had a weakly positive DAT the cord serum bilirubin concentration never exceeded 3.4 mg/dl (58 µmol/l) and no infant developed hyperbilirubinaemia severe enough to require phototherapy (Bowman et al. 1978). In a few cases in which D-negative women were given four doses of 300 µg of anti-D between about the twelfth and thirty-fourth weeks of pregnancy in an attempt to turn off the immune response following the discovery of low concentrations of maternal anti-D reacting only with enzyme-treated cells, not all the D-positive infants born subsequently had a positive DAT (JM Bowman, personal communication). Antenatal administration of anti-D is reported to effect a rise in plasma levels of cytokines TGF-beta and prostaglandin E2 (Branch et al. 2006). These authors speculate that elevated levels of these cytokines are relevant to the mechanism of immune suppression induced by anti-D. In a predominantly white population about 38% of women receiving anti-D will be carrying a D-negative fetus and so receiving anti-D unnecessarily. Routine ante-natal D typing of fetal DNA from maternal plasma could avoid this unnecessary exposure (see above).
Results Observed with Antenatal Treatment
A review of 11 published trials (seven non-randomized trials with historical or geographical controls, one randomized control trial, one quasi-randomized controlled trial, one community intervention trial and one retrospective before-and-after study) concluded that routine antenatal anti-D prophylaxis is effective in reducing the number of D-negative women who are sensitized during pregnancy (Chilcott et al. 2003). These authors suggest that introduction of routine antenatal anti-D prophylaxis in England and Wales would reduce the sensitization rate from 0.95% to 0.35%. Furthermore, such an intervention is cost-effective, at least in the UK (Chilcott et al. 2004). In one series in which almost 10 000 D-negative women carrying D-positive fetuses were given an injection of anti-D at 28 weeks (either 300 µg i.m. or 240–300 µg i.v.), less than 0.1% developed anti-D by full term, whereas previous experience indicated that without antenatal anti-D the incidence would have been 1.8% (Bowman 1984). As explained in ‘Anti-D present by the end of a first full-term pregnancy’, the figure of 1.8% is substantially greater than that reported by others, but the discrepancy can be explained.
In another series in which, unlike the one just referred to, only primiparae were included, of 1238 D-negative women carrying D-positive fetuses who were injected at both 28 and 34 weeks with 100 µg of anti-D, only 0.16% developed anti-D by the time of delivery. In an earlier series from the same centre in which women were given anti-D only after delivery, 0.9% formed anti-D by the end of pregnancy (Tovey et al. 1983). The foregoing figures give a slightly too favourable impression of the success of antenatal immunoprophylaxis with anti-D because in both series a few additional women developed anti-D either 6 months after delivery or during a second pregnancy. In the first series, in which most of the women were tested at 6 months, there were two in whom anti-D was detected for the first time, an incidence of very approximately 0.02%; of an unknown number who had a second D-positive pregnancy there were another two who developed anti-D before the twenty-eighth week (Bowman 1984). In the second of the two series quoted above anti-D developed during a second pregnancy with a D-positive infant in 2 out of 325 women, but in neither case did the antibody appear to cause clinically significant haemolytic disease. New anti-D immunizations were reduced from 0.67% to 0.31% by a single injection of 200 µg anti-D in the 30th week of pregnancy in a study from the Netherlands (Koelewijn et al. 2008).
Amount of Anti-D for Antenatal Injection
From the data of Bowman and co-workers (1978) it appears that treatment should be given as early as 28 weeks. In the USA and some other countries, the only antenatal dose of anti-D is 300 µg, injected at 28 weeks. In nine women who had received this dose, plasma levels of anti-D 9–10 weeks later were between 0.7 and 2.6 ng/ml, corresponding to a total residue of 6–21 µg of anti-D. At term, some women had no detectable anti-D (Bowman and Pollock 1987). Better survival of injected IgG anti-D, indicating that an average of 19 µg should remain 12 weeks after a dose of 250 µg, was found in another investigation, although there was substantial variation between different women (Eklund et al. 1982). It must be concluded that a single dose of 300 µg given at 28 weeks may be too small to protect all women (Bowman and Pollock 1987). The administration of 300 µg of anti-D at 28 weeks is convenient in countries where 300 µg of or thereabouts is the standard dose to be given postpartum, as the same preparation can then be given on the two occasions. On the other hand, in countries where the standard postpartum dose is 100 µg it is convenient to give two doses of 100 µg during pregnancy, at 28 weeks and 34 weeks respectively. This dosage will in fact result in a rather higher concentration of anti-D in the mother’s plasma immediately before delivery than when a single dose of 300 µg is given at 28 weeks (seventh edition, p. 395), will result in a certain saving of anti-D and, again, will avoid the confusion of having two different sized doses to be administered antenatally and postnatally.
When anti-D is given antenatally, the serum concentration at term may still be high enough to be detectable by the IAT. In women who have been given anti-D Ig antenatally, the finding of a positive IAT at the time of delivery must not be regarded as a contraindication to giving the usual postpartum dose.
Intravenous Administration of Anti-D
As discussed in Chapter 5, it is possible that when anti-D is given intravenously at the same time as an injection of D-positive red cells, the dose of antibody required for the suppression of primary Rh D immunization may be smaller than when the anti-D is injected intramuscularly. There is therefore a potential advantage in giving anti-D intravenously rather than intramuscularly immediately following delivery when it is likely that any D-positive red cells in the circulation have arrived there only recently. Intravenous anti-D was used successfully to clear a massive fetomaternal haemorrhage (142 ml) with a t1/2 of 24.5 h by Larsen et al. (2008). The mother had no detectable anti-D six months after delivery. In practice, because of the increased risk of transmitting viral infections when Ig is given intravenously (see Chapter 16) and because the methods used for viral inactivation in some preparations are suspect, the i.m. route should be used whenever possible. When giving anti-D antenatally there does not seem to be any potential advantage in i.v. administration, as the object is to protect against Rh D immunization over a period of many weeks and, within 3 days of giving anti-D, plasma levels are the same whether the injection has been given intravenously or intramuscularly.
Occasional reactions have been reported following the i.v. injection of anti-D immunoglobulin prepared by fractionation on DEAE Sephadex (Hoppe et al. 1973); at one centre, only seven reactions, two of which were severe enough to require treatment, were noted following injections to 80 000 women (H Hoppe, personal communication). In another series, transient marked flushing and mild chest discomfort were noted in 2 out of 2792 women. The reactions were thought to be due to aggregated material in vials that had too high a moisture content (Bowman et al. 1980). In a third series, not a single untoward reaction was reported amongst 120 000 recipients (JR O’Riordan, personal communication). In a fourth series involving 261 women no adverse reactions were reported (MacKenzie et al. 2004).
Monoclonal/Recombinant Anti-D for Prophylaxis
Although human monoclonal anti-D derived from Epstein–Barr virus (EBV)-transformed B lymphocytes has been available since 1984 (Crawford et al. 1983), it has not yet emerged as a licensed product capable of replacing the tried and tested polyclonal product described above. Two monoclonal antibodies, one IgG1 (BRAD5) and one IgG3 (BRAD3), have been used as a blend in trials and shown to be capable of protecting D-negative male volunteers from D immunization (Kumpel et al. 1995; Smith et al. 2000). BRAD5 can be quantified in human serum using a monoclonal anti-BRAD5 idiotype by ELISA (Austin et al. 2001). Monoclonal anti-D produced by EBV transformation of B lymphocytes can be manipulated using recombinant DNA technology to optimize affinity and Fc effector functions (Proulx et al. 2002) and expressed in alternative expression systems for manufacture (Edelman et al. 1997; Miescher et al. 2000). A recombinant IgG1 anti-D produced in CHO cells was shown to prevent D immunization in male volunteers (Miescher et al. 2004).
Recombinant Mutant Anti-D for Treatment of Severe Cases of HDFN
It is possible to engineer recombinant anti-D lacking effector functions (Armour et al. 2006; Nielsen et al. 2008) raising the possibility that inactive mutant anti-D could be used as a therapy in situations where the fetus is at risk of severe HDFN in utero. In this scenario mutant anti-D given to the mother crosses the placenta and coats fetal red cells, thereby preventing binding and consequent red cell destruction resulting from binding of functional anti-D. There are however, formidable problems to be overcome before such a therapy could become a reality (reviewed by Urbaniak 2008). These are most notably, identifying the effective dose of antibody, the number of doses required during the pregnancy and surmounting the regulatory hurdles to allow clinical trial of such a product in pregnant healthy women.
Immunosuppression of D Sensitization by Induction of Tolerance
Successful induction of tolerance to peptides corresponding to regions of the D polypeptide in transgenic mice presages the development of a new approach to prevention of haemolytic disease resulting from D immunization (Hall et al. 2005; see also Chapter 3).
Paternal Leucocyte Therapy
Lam and co-workers (2003) investigated the feasibility of isolating paternal mononuclear cells free of red cell contamination with a view to using these cells as a treatment for severe D alloimmunization.
Changes in the Incidence and Mortality of Haemolytic Disease
As described in Chapter 3, before the introduction of immunoprophylaxis of Rh D haemolytic disease, the frequency of the disease, both in England and in North America was about 1 per 170 births. Assuming an average family size of more than two children, this figure is close to expectation: the frequency of the disease in first pregnancies is very low (less than one per 1000 births), in second pregnancies is about one per 100 births, and in subsequent pregnancies is somewhat greater.
Immunoprophylaxis became available in 1968–70. In one North American hospital (dealing with a predominantly white population), in which postnatal anti-D was introduced in 1968, and antenatal anti-D added in 1985, the frequency with which (immune) anti-D was found in pregnant women declined from 1 in 238 in 1974 (Walker 1984) to 1 in 963 in 1988 (Walker and Hartrick 1991) and to 1 in 1663 in 1990–92; in the last period, HDN was diagnosed in only 1 in 2419 births (Walker et al. 1993).
In one English region (East Anglia), the figures for 1993–95 were as follows: anti-D was found in one per 584 pregnant women but in almost 80% of cases the anti-D was not detected at the time of the first antenatal visit and was subsequently present at a concentration of less than 1 iu/ml (0.2 µg/ml). Less than one infant per 2000 had clinical signs of HDN (A Rankin, personal communication). It should be noted that anti-D is given antenatally in only approximately 12% of hospitals in England and Wales (Chilcott et al. 2003).
The decline in the death rate from Rh haemolytic disease has been substantially greater than the decline in the incidence of the disease. In England and Wales, registered deaths fell from 1 in 2180 births in 1953 (Walker 1958) to 1 in 5400 in 1977 and to 1 in 62 500 in 1990 (Clarke and Mollison 1989; Hussey and Clarke 1992). It must be emphasized that these figures are not for total deaths, as up to September 1992 deaths occurring before the twenty-eighth week of pregnancy were not registered in England and Wales. (Since October 1992, deaths from 24 weeks onward have been registered.) Chilcott and coworkers (2003) put the figure at one death in every 20 800 births.
The Most Important Steps in Preventing Rh D Immunization by Pregnancy
First, in order to minimize the risk that, due to a technical or clerical error, a D-negative woman will be reported as D positive and thus not be given anti-D immunoglobulin after delivery, the D group of all so far unimmunized women should be determined (using two different anti-Ds) on at least two separate occasions: (1) at the time of the first antenatal visit (booking); and (2) at 28–34 weeks. In addition, the following steps are recommended for all so far unimmunized D-negative women:
Related posts:
 Immunology of leucocytes, platelets and plasma components
Immunology of leucocytes, platelets and plasma components
 Red cell antibodies against self-antigens, bound antigens and induced antigens
Red cell antibodies against self-antigens, bound antigens and induced antigens
 ABO, H, LE, P1PK, GLOB, I and FORS blood group systems
ABO, H, LE, P1PK, GLOB, I and FORS blood group systems
 Alternatives to allogeneic transfusion
Alternatives to allogeneic transfusion
 The transfusion of platelets, leucocytes, haematopoietic progenitor cells and plasma components
The transfusion of platelets, leucocytes, haematopoietic progenitor cells and plasma components
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


