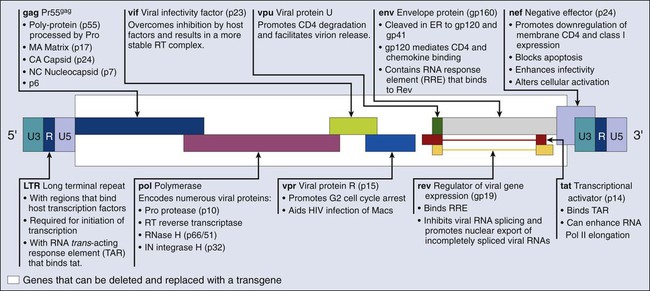
James E. Talmadge and Kenneth H. Cowan Recent Advances in Gene Therapy • Chimeric antigen receptor transduction of peripheral blood T lymphocytes for delivery of specifically cytotoxic immunoreactivity • Silencing of gene expression with short hairpin RNA delivered with nanoparticle or lentiviral delivery vectors • Regulated transgene expression • Can be targeted either physically or via promoter expression and is nontoxic, noninflammatory, and nonimmunogenic • Should have the potential to incorporate a large transgene and result in high levels of both transduction and transgene expression • Duration of transgene expression and/or genomic integration ought to be regulatable Current Concerns Regarding Gene Therapy • Gene therapy using an adenovirus vector has been implicated in the death of at least one patient • Leukemic transformation by insertional mutagenesis Future Directions of Gene Therapy • The choice of disease, clinical implementation, and vector are critically important to the future development of successful gene therapy • Because of deficiencies in gene delivery and targeting, as well as expression levels, it is critical to pair protocols with specific vector attributes In 2003, the first gene therapeutic agent was approved in China. This agent was an Adv serotype 5 vector engineered to express TP53 (Gendicine) for treatment of patients with head and neck squamous cell carcinoma (HNSCC).1 A second gene therapy product, H101 (ONYX-015), an Adv vector modified to replicate in and kill cancer cells with TP53 mutations, was approved in December 2005.2 More recently (in December 2011), Neovasculogen (a plasmid vector with a vascular endothelial growth factor [VEGF] transgene) was approved in Russia for the treatment of peripheral arterial disease. Regardless of these approvals, the primary challenges in gene therapy remain improvements in the targeting of our existing vectors and increasing gene transduction efficiency. Overcoming these obstacles will facilitate the development of targetable vectors, and given the systemic nature of most malignancies, will help in the development of vectors that can be administered intravenously. This chapter focuses on strategies to improve efficacy and reviews ongoing gene therapeutic strategies. Recent advances are also examined and discussed, and areas are indicated that require further development for clinical gene therapy to become a widely used treatment modality. Viral gene delivery has developed from the innate ability of a virus to infect T cells, which offers many intrinsic advantages3,4: • Specific cell-binding and cell-entry properties • Efficient targeting of the transgene to the nucleus of the cell The Retroviridae is a large family of RNA viruses including Moloney-murine-lentivirus–related viruses (e.g., Moloney murine leukemia virus [MLV]) and lentiviruses (e.g., human immunodeficiency virus type 1 [HIV-1] and HIV-2).5 Their genomes consist of two identical positive-sense, single-stranded RNA molecules (~3.5 kb) and are encased in a capsid along with integrase and reverse transcriptase enzymes. Initially, retroviral vectors were the most widely used viral vectors, a distinction that has been replaced by Adv vectors in recent years. Retroviruses can transduce only those cells that are actively undergoing mitosis, limiting their utility with certain cell populations, especially hematopoietic stem cells (HSCs). Retroviral vectors provide good gene expression and are technically easy to produce, although the titers obtained are suboptimal. In addition, the production of retroviral vectors needs to be carefully monitored because of the potential for helper virus contamination. Most of the retroviral vectors that are used for gene therapy are based on the Moloney MLV. Vector replication is prevented by the deletion of the gag, pol, and env gene regions. The gag region encodes the capsid proteins; the pol region encodes reverse transcriptase and integrase; and the env region encodes proteins required for receptor recognition and envelope anchoring (Fig. 31-1). The genome includes long terminal repeats at either end that play a vital role in initiating DNA synthesis and regulating transcription of the viral genes. The gag, pol, and env gene products are supplied by a complementary packaging cell line. When a retroviral vector plasmid is introduced into a packaging cell line, viral RNA is produced, packaged into virions, and secreted into the medium. Each resultant viral particle is able to integrate itself into the genome of the host cell but is unable to produce additional viral particles because it lacks the gag, pol, and env genes. The transduced DNA sequences are stably integrated into the chromosomal DNA of the target T cells and in this way are transferred to cellular progeny of transduced cells. Highlights of results obtained to date with retroviral vectors include therapeutic studies in children with severe combined immune deficiency (SCID-X1), which will be discussed later in this review, as well as the gene-marking studies of Malcolm Brenner and others.6,7 In the latter studies, it was shown that tumor cells within autologous stem cell transplant products could be responsible for tumor relapse, at least in patients with leukemia. The most recently discovered members of the retrovirus family are the human and simian immunodeficiency viruses, which belong to a subclass of retroviruses known as lentiviruses.8,9 The development of HIV gene therapy vectors has several potential advantages based on the following characteristics: • Transduction of actively dividing and nondividing cells • Long-term, stable transgene expression as a result of genetic integration Genetic modifications, such as the introduction of vesicular stomatitis virus (VSV) G protein into the lentiviral envelope, can widen the tropism of this vector. The first clinical study using a lentivirus vector was undertaken in HIV-infected patients.10 This study investigated the safety of infusing autologous T cells modified with an HIV-1–based lentiviral vector expressing an antisense gene against the HIV envelope. Five patients with HIV infections that were resistant to antiviral therapy and who had viral loads of >5000 copies/mL and CD4+ T-cell counts between 200-500 cells/mm3 were treated. The primary end points included adverse events, viral load, CD4+ counts, and the emergence of replication-competent lentivirus derived from the vector. In this phase I study, one subject was reported to have a sustained decrease in viral load. The CD4 counts remained steady or increased in the other four subjects, and sustained gene expression was observed. These preliminary studies support the safety of lentivirus vectors. Recombinant Adv is a nonenveloped, icosahedral, double-stranded DNA virus with a capsid containing 252 capsomeres (240 hexons and 12 pentons).11 The large genome of Adv (36 kb) enables large genes to be inserted into an Adv-based vector. Transgenes in Adv vectors are not incorporated into the genome of transduced cells, but rather remain as an extra chromosomal entity in the nucleus. First isolated from U.S. Army recruits who had acute respiratory symptoms, Adv vectors have been found to be common human pathogens. To date, 49 serotypes have been characterized and associated with a variety of symptoms, ranging from a mild cold to acute febrile pharyngitis. Replication-defective recombinant Adv vectors are currently the most commonly used viral vectors in clinical trials. Ad2 and Ad5 are used primarily for gene therapy applications. Recently, however, the Ad11 and Ad35 serotypes were shown to exhibit a unique tropism that includes HSCs, a finding that potentially widens their utility.14–14 The Adv vector’s genome (Fig. 31-2) can be divided into two main regions: early (E) and late (L), according to the time at which their genes are expressed during virus replication. Four regions of early genes are termed E1, E2, E3, and E4, and one region of late genes comprises the five coding units termed L1, L2, L3, L4, and L5. The E1 region is essential for viral replication; therefore recombinant Advs without the E1 region are considered replication defective. In a replication-defective Adv vector, the E1 region can be replaced with a transgene for expression. Further, removal of genetic material from the vector, such as the E3 and/or the E4 region(s), allows for larger genes to be inserted and reduces the viral immunogenicity.15 Viruses without the E3 and E4 regions are referred to as “gutless” and have decreased antigenicity.16 The late genes (L1 to L5) are expressed at the onset of viral DNA replication and code for structural polypeptides that are needed for virion assembly. This understanding of viral replication has allowed the development of extremely elaborate, conditionally replicative Adv vectors capable of replication only in cancer cells.17 However, Adv vectors have potential shortcomings, including: • Transient expression because the viral DNA does not integrate into the host • Viral protein expression by the Adv vector after administration into a host • Adv is a common pathogen, resulting in hampered in vivo delivery associated with antibody responses4 Because the period of Adv transgene expression is relatively short, this vector is suboptimal if expression is desired for longer than 10 to 14 days. This short expression time is due primarily to the induction of a cytotoxic T lymphocyte (CTL) response to viral polypeptides, as well as potentially to the transgene itself, especially if it is not expressed normally. Because the Adv genome does not integrate into the target cell, only one of the daughter cells (if the target T cells are dividing) will contain the transgene. Manipulation of the immune response can result in longer expression; however, Adv gene delivery is ideally suited to situations that require only a single period of transgene expression in which transient expression is desired—for example, growth factor therapy. A second major disadvantage of Adv vectors used in vivo is the immune response (CTL and antibody [Ab]), both endogenous and induced, which can preclude infection and cause the destruction of transduced cells, resulting in local tissue damage and inflammation. This shortcoming was demonstrated in studies with intrabronchial delivery of Adv for the treatment of cystic fibrosis.18 Host T cells presenting peptides from Adv-encoded transgene products target the host cell for CTL-mediated destruction. A third major disadvantage of Adv vectors is that most humans are primed against at least one serotype, because Adv is a naturally occurring virus. Using the same serotype in a gene therapy context will likely result in a rapid and vigorous immune response, such that high levels of anti-Adv Ab occur in the sera within days of Adv vector administration. Another similar problem is the potential secondary immune response induced by the readministration of a vector. It must be stressed that transgene expression can occur during a boost, although a shortened duration is observed. The augmentation of a CTL response by an Adv vector suggests the utility of Adv vectors as vaccine adjuvants (Box 31-1). Adeno-associated virus (AAV) vectors offer many of the same advantages as Adv vectors, including a wide host-cell range and a relatively high transduction efficiency.19,20 AAV vectors stably integrate at specific sites in the host genome, resulting in a longer lasting transgene expression. In addition, these stable vectors can infect a variety of dividing and nondividing cells without inducing an immune response. AAV vectors cause little damage to target T cells, unlike Adv vectors, which can cause a high degree of cytopathogenicity. However, evidence suggests that AAV vectors are significantly less efficient than retroviral vectors at transducing primary cells, because most of their DNA remains extrachromosomal and does not integrate into the host genome. Furthermore, they cannot incorporate genes larger than 5 kb and must be screened closely for Adv contamination. Herpes simplex virus (HSV) vectors are developed primarily for protocols that target neuronal tissue. Similar to Adv vectors, HSV vectors are maintained as an extrachromosomal DNA element in the nucleus of host T cells but can establish long-lived asymptomatic infections in the sensory neurons of the peripheral and central nervous tissue.21 HSV vectors also have a wide host range and are similar to Adv vectors in that they allow large gene inserts of up to 20 kb. These vectors are infective even with multiple deletions of immediate-early genes that are essential for replication, resulting in less cytotoxic vectors, thereby reducing safety concerns.22 HSV vectors can be produced at high titers and express transgenes for a long period in the central nervous system.23 The major concern associated with HSV is the potential for wt virus to replicate lytically in the human brain, resulting in encephalitis. Other significant disadvantages with HSV vectors include: The origin of vaccinia virus (VV), the virus used for vaccination against smallpox, is not known, but it was probably derived from cowpox virus, variola virus, or a hybrid of the two.24,25 Percutaneous VV vaccine administration results in protective cellular and humoral immune responses in more than 95% of primary vaccinees. Recombinant VV vectors are highly attenuated, host-restricted, and nonreplicating or poorly replicating poxvirus strains (including the modified vaccinia Ankara [MVA] and canarypox or avipox vector [Alvac]) and thus do not create productive infections.26,27 MVA is avirulent in normal and immunosuppressed animals and safe in humans.28 Recent studies using transgenic mice provided a comparison of VV immunogenicity, including MVA and Western Reserve (WR). These studies demonstrated that MVA vaccines elicited CD8+ T-cell responses that are comparable with those induced by the replication-competent WR strain. Further, MVA vaccination was shown to be protective against a lethal respiratory challenge with the virulent WR strain.29 The most frequent adverse complication of VV vaccination is inadvertent inoculation (usually autoinoculation) at other sites. Serious complications, which are more common among primary vaccinees and infants than among revaccinees and adults, include the following: • Generalized vaccinia in otherwise healthy individuals, which is generally self-limiting • Eczema vaccinatum, which consists of disseminated cutaneous lesions in highly susceptible patients with eczema or other chronic skin diseases and can be severe or even fatal • Progressive vaccinia (vaccinia necrosum), which is a severe, potentially fatal illness seen in patients with immunodeficiency, whether congenital, acquired (e.g., via leukemia or lymphoma), iatrogenic (e.g., via chemotherapy or glucocorticoid treatment), or induced by HIV • Postinfectious encephalitis, which is rare (three cases per million primary vaccinees) but can be fatal in 15% to 25% of cases and can leave 25% of patients with permanent neurological sequelae Similar to Adv vectors, VV vectors are used for immune manipulation and as a vector for vaccines.30 VV vectors have been used worldwide to eradicate smallpox and, as discussed previously, to provide a relatively safe live vaccine. Vaccinia vectors do not integrate into the genome of the host cell; however, they can accommodate large transgenes and are extremely immunogenic. VV vectors are used to immunize patients against tumor antigens by cloning antigens and/or genes encoding proteins with adjuvant activity (e.g., cytokine or costimulating factor genes) into the viral genome. Most transgenes are expressed at high levels in vivo, eliciting an antigen-specific response. Vector-induced immunity, however, can limit the ability of the vaccinia transgenes to boost an immune response, which is an observation similar to that seen with Adv vectors. The current emphasis is on VV infection of dendritic cells (DCs) using a vector with an antigenic transgene.30,31 In association with the immunogenicity of VV vectors and their ability to deliver an antigenic transgene, they have been used clinically as a melanoma vaccine. In clinical studies by Wallack and colleagues,32 a phase III trial of a vaccinia melanoma oncolysate, delivered as an active specific immunotherapy, was found to increase the disease-free or overall survival (OS) of patients with stage III melanoma in a surgical adjuvant setting. Other studies have used VV mutants that are conditionally replicative and can lyse cancer cells after viral replication. These vectors have been used in a strategy whereby insertional inactivation of the VV thymidine kinase (TK) gene was used to limit viral replication in cells with large intracellular nucleotide pools, such as tumor cells. In a similar approach, Mastrangelo and coworkers33 inserted the gene for granulocyte macrophage-colony stimulating factor into the VV TK gene locus as a strategy to generate an oncolytic virus that induced antitumor immunity after infection of malignant melanoma. This vector is currently in a clinical trial of intralesional administration to patients with refractory recurrent melanoma. In the first seven patients studied, two patients had a complete response and three other patients had partial responses. Other oncolytic VV vectors have been engineered with complementary DNAs for cytokines such as interleukin 2 (IL-2) or with pro–drug-activating enzymes such as cytosine deaminase (CD) to augment antineoplastic efficacy.34,35 The role of VV vectors as vaccines has focused predominantly on carcinoembryonic antigen (CEA) as the vaccine antigen. CEA is a glycoprotein self-antigen found in breast, lung, gastric, colon, and ovarian tumors. One such vector is a recombinant VV containing the CEA gene (rV-CEA).36,37 In a phase 1 clinical trial, the safety of rV-CEA was demonstrated; however, no significant antineoplastic effects were observed.38 Possible reasons for the lack of clinical efficacy in these trials include: • Prior exposure to the VV, leading to the development of antivaccinia immune responses after repeated vaccinations • Advanced state of the patients’ tumors Another phase 1 rV-CEA vaccine study demonstrated that CEA-specific T-cell responses could be generated in humans after vaccination.38 A second recombinant anti-CEA vaccine, Alvac-CEA, has been developed.39,40 As with rV-CEA, Alvac-CEA contains the CEA gene; however, unlike rV-CEA, it cannot replicate in mammalian cells. The safety of Alvac-CEA has been documented in phase 1 trials in patients with advanced carcinomas.41 A moderate but statistically significant increase in the number of CEA-specific CTL precursors was observed in seven of nine human leukocyte antigen (HLA)-A2–positive patients treated with Alvac-CEA, although objective anticancer effects were not observed. Preclinical studies have suggested that the combination of rV-CEA and Alvac-CEA in a prime and boost protocol can induce a more vigorous T-cell response than either vaccine alone.39 In a clinical prime and boost study, 18 patients with advanced tumors expressing CEA were randomly assigned to receive either rV-CEA followed by three Alvac-CEA vaccinations, or Alvac-CEA (three times) followed by one rV-CEA vaccination. In this study, vaccination with rV-CEA followed by Alvac-CEA resulted in an increased frequency of antigen-specific interferon gamma (IFN-γ+) cells by enzyme-linked immunospot assay relative to the reverse order of vaccination.42 Another method to enhance the responses to a vaccine is to incorporate a costimulatory signal. In the absence of a costimulatory signal, presentation of an antigen to T cells can result in anergy.43 B7.1, which binds to CD28 on T cells, is one such costimulatory signal that results in the production of IL-2 and IFN-γ by T cells. In a vaccine study using VV vectors, 39 patients were treated with Alvac-CEA B7.1.44 In one study using the Alvac-CEA-B7.1 vaccine, patients with metastatic CEA-expressing adenocarcinomas received vaccine intradermally every 2 weeks for a total of four injections. In this phase 1 trial, 27% of the patients had disease stabilization after four vaccinations. Six of 31 patients with elevated serum CEA levels had a temporary decline in CEA. In addition, HLA-A2–positive patients demonstrated increased CEA-specific T-cell frequencies after three vaccinations. Based on these studies and additional phase 2 data, a phase 3 trial was initiated in 255 patients with advanced pancreatic cancer at approximately 60 medical centers.45 The protocol was powered to detect a 2-month improvement over control chemotherapy based on a median OS of 6 months. Unfortunately, this study did not meet its primary end point of improving OS compared with palliative chemotherapy or best supportive care. However, this outcome is not unexpected, because patients with advanced pancreatic cancer have a rapid disease progression and are poorly responsive to intervention in general (Box 31-2). High-titer alphavirus vectors can provide efficient gene delivery both in vitro and in vivo. In addition, efficient central nervous system infections via intranasal and vascular injections with virulent and avirulent replication-competent Semliki Forest virus (SFV) strains have been shown in animal models.48–48 Replication-deficient alphavirus particles have a high local and transient transgene expression in rodent brains. Further, repeated SFV injections are possible in the absence of an immunogenic response against SFV, which is in contrast to Adv and VV vectors. Modifications to the envelope structure of Sindbis virus are possible with resultant changes in host range and targeting. The favorable characteristics of alphavirus vectors include: Nonessential genes can be removed from viral vectors to allow room for transgene(s) to reduce inflammatory responses and to increase safety.49,50 This process involves simplifying the virus, sometimes to an extreme. After undergoing such a process, a virus vector can be an artificial “vector shell,” allowing the gene of interest to be expressed at high levels, in a highly regulated manner, and for a controlled period. Another approach to achieve the same result is to produce a vector that can introduce genetic material to the nucleus of cells.49,51,52 This strategy has resulted in the development of several nonviral vector systems; however, the efficiency of “naked DNA” as a therapeutic is suboptimal without some form of carrier or formulation. One form of nonviral gene delivery is the use of purified DNA plasmids. The transgene expression is low after intramuscular or intratumoral (IT) injection; however, high levels are observed if hydrodynamic injection is employed.53,54 The approach of naked DNA injection is typically done as an intramuscular or IT injection. Despite the simplicity of this approach, transfection efficiency is low and results in limited expression. Various formulations, including lipid or pluronic formulations, and incorporation into nanoparticles or liposomes have been used to improve transduction efficacy and gene expression (Box 31-3).57–57 Nonviral liposomal delivery systems can be intravenously injected with limited vector-associated toxicity, but with transgene expression, especially in the lungs.58 Tumor targeting using tumor-specific promoters, ligandation of receptors to the liposome surface, and PEGylation of liposomes have all been studied.59–65 Although some degree of tumor targeting has been observed using these delivery systems, the level of transgene expression is generally low. Studies have revealed that liposome-DNA complexes can also elicit an inflammatory response when injected systemically, resulting in suppression of transgene expression.68–68 Furthermore, failure to achieve increased or sustained gene expression after repeated injections has been a major obstacle in the development of liposomes.67,69 Recently, it was shown that cationic liposome (DOTAP : cholesterol or DOTAP : Chol) DNA complexes can achieve effective levels of transgene expression in tumor-bearing lungs, and when injected intravenously, can achieve levels sufficient to cure immunocompetent mice with disseminated experimental metastases.70 Further, repeated daily injections can result in a dose-dependent increase in transgene expression in tumor-bearing lungs.71 Hydrodynamic tail vein plasmid delivery results in high levels of transgene expression in the livers of rodents.72 Lower levels of transgene expression (100- to 1000-fold) are found in the spleen, heart, kidneys, and lungs. This simple nonviral gene transfer procedure entails the rapid delivery of naked plasmid DNA in a relatively large volume of physiological saline solution.53 In a typical mouse weighing 20 g, the plasmid is delivered in a total volume of 2.0 mL over a period of 5 to 7 minutes. In their most basic form, liposomes consist of two lipid species: a cationic amphiphile and a neutral phospholipid.71,73 Liposomes spontaneously bind to and condense DNA to form complexes that have a high affinity for the plasma membranes of cells, resulting in the uptake of liposomes to the cytoplasm by endocytosis. Many variations of this approach are used, resulting in varying levels of gene expression. Unfortunately, liposome-facilitated gene delivery is relatively ineffectual in vivo. More recently, some of the advantages of viral delivery vectors have been combined with the safety and “simplicity” of the liposome to produce fusogenic virosomes.73 Virosomes are engineered by complexing the membrane fusion proteins with liposomes that have already encapsulated plasmid DNA. The inherent ability of the viral proteins in virosomes to fuse with cell membranes results in the efficient introduction of DNA to the target cell, providing improved gene expression. Viral vectors have limitations based on the size of transgene that can be incorporated; in contrast, no such limit exists for virosome or liposome technology (at least in theory). The ballistic physical method of gene delivery uses microcarriers (usually gold particles) coated with DNA and “fired” using an explosive or gas-powered ballistic device called a “gene gun.”76–76 Once the particles are inside the target cell, the DNA is slowly released from the microcarriers, transcribed, and translated. This application has been used extensively in animal models, but its clinical use is restricted to exposable surfaces or ex vivo transduction because the particles do not penetrate tissues deeply.77 Novel polymeric delivery systems (e.g., nanospheres) are potentially useful because the smaller the size of the condensed DNA particles, the better the in vivo diffusion to target T cells and trafficking within cells.80–80 Individual plasmid molecules can be condensed into a nanoparticle using detergents. Thus nanoparticle-based gene delivery targeting the neovasculature using an integrin-targeting ligand has been shown to result in tumor regression.81 Nonetheless, the size of the transgene that can be delivered is limited, and the primary focus has been on small interfering RNA (siRNA) delivery, which is discussed in a later section. High molecular weight, double-stranded DNA constructs containing transgenes are frequently used for gene therapy to introduce transgenes into cells that inherently lack the ability to produce a protein of interest. In addition to being used in the treatment of congenital diseases, DNA vectors are also used as vaccines for genetic immunization.82 Suicide gene therapy is another rapidly emerging strategy in which chemosensitization genes are delivered to tumor cells; upon gene expression, they convert a separately administered, nontoxic prodrug into a chemotoxic drug.82,83 Because only the transfected tumor cells can convert the prodrug, the susceptibility to the chemotoxic activity is limited to the tumor cells, hence the term “suicide gene therapy.” To date, the primary transgene source for DC transduction is DNA, although other antigen sources are also used with DCs, including peptides, recombinant or purified proteins, cellular extracts from tumor cells, apoptotic bodies, and RNA or DNA plasmid vectors. Nevertheless, the carrier of choice for loading DCs with tumor antigens is DNA or RNA.84 Nucleic acid transfection leads to the display of multiple antigenic epitopes by both classes I and II major histocompatibility complex via the antigen-processing machinery of the patients’ DCs, resulting in the display of the “most appropriate” peptides. This strategy contrasts with vaccine strategies based on synthetic peptides, which require the knowledge of the patient’s unique peptide epitopes. Thus nucleic acid transfection of DCs offers several advantages for both immunologic and practical considerations. The bias for the use of DNA vectors includes an increased stability compared with RNA, the ability to produce plasmids in large quantities, and the ease with which the sequence can be modified to regulate expression.85 In several respects, however, RNA vectors are also advantageous when compared with DNA transfection.85 RNA vector advantages include the ability to use total messenger RNA (mRNA) isolated from tumors to transfect DCs with no intervening cloning steps and the ability to express several or potentially all tumor-derived genes within DCs. Transfected RNA need only reach the cytoplasm of DCs, whereas DNA requires entry into the nucleus and subsequent transcription. Thus the low levels of antigenic epitope expression with RNA-transfected DCs could be advantageous, provided that expression levels are sufficient to generate a T-cell response.84 The low levels of antigenic peptides presented by DCs allow only the T cells with high-affinity receptors to be activated, skewing the response toward T cells better able to recognize the tumor cells. Conversely, DCs presenting high levels of antigenic peptides allow T cells with low affinity receptors to be activated, thus masking or even preventing response of high-affinity T cells. This could result in T cells that kill cells with high antigen expression, but cannot kill tumor cells, which typically express low antigen levels. Thus RNA-transfected DCs potentially have greater efficacy for the activation of high-affinity T cells.85 Oligonucleotides are short single-stranded segments of DNA or RNA that, upon cellular internalization, can selectively inhibit the expression of a single protein.86 Multiple forms of oligonucleotides are used in gene therapy, including antisense, siRNA, and ribozymes. Most of these constructs form a duplex with the mRNA or the pre-mRNA and inhibit their translation or processing, consequently inhibiting protein biosynthesis. This process occurs by multiple mechanisms, as discussed in the following sections. RNA interference is a specific gene-silencing mechanism induced by the delivery of synthesized siRNA. siRNA is generated by dicer, an endonuclease that cleaves long double-stranded RNA molecules into fragments of 21 to 23 base pairs, which are highly specific for the nucleotide sequence of its target mRNA.87–90 These siRNAs associate with helicase and nuclease molecules and form a large complex, termed “RNA-induced silencing complex,” that unwinds siRNA and directs sequence-specific degradation of mRNA. RNA interference is a highly conserved molecular mechanism used by eukaryotic organisms to control gene expression and to defend their genomes against invaders, such as transposons or RNA viruses. siRNA-based technology has the potential to develop into an effective therapeutic strategy because it can modulate the expression of any protein, even undruggable target proteins such as transcription factors, with high specificity. However, the clinical development of siRNAs has been slow because of limited cellular uptake, low biological stability, and suboptimal pharmacokinetics. Indeed, naked siRNAs are easily degraded by nucleases and rapidly eliminated, and their negative charge and hydrophilic nature impairs cellular internalization.91 Advances have been achieved with lipid-based nanocarriers that are sterically stabilized by polyethylene glycol (PEG); however, PEGylated liposomes have technical challenges for siRNA delivery, and exchangeable PEG-derivatized lipids have been developed to improve tumor gene expression.92 Currently, lentiviral vectors provide a good option for siRNA delivery to cells, including nondividing cells.93 Recently it has been shown that siRNA is active in vivo, resulting in therapeutic activity.94 In one study, siRNA knockdown of the mutant K-ras oncogene had pronounced antitumor activity.95 In this study, siRNA was delivered as a nonreplicative retroviral transgene and was shown to inhibit the relevant mutant K-ras and prevent anchor-independent growth and tumorigenicity. Despite these advances, RNA interference technology still requires refinement before its full potential can be utilized for routine clinical treatment. Antitumor activities can also be obtained through siRNA knockdown of other critical components of tumor growth, metastasis, angiogenesis, and chemoresistance.96 Stable transfection and expression of siRNA occurs with nonreplicating viruses97,98; however, oncolytic virus vectors provide another method to extend bioactivity. This tumor-selective infectivity can restrict transgene expression to the cancer microenvironment, potentially reducing toxicity and extending transgene expression via viral replication and multiple cycles of infection within permissive cancer cells.99,100 Furthermore, viral oncolysis can augment antitumor outcomes by siRNA-mediated therapeutic activity. In one study,101 the replication-competent oncolytic Adv ONYX-411 was used to deliver a mutant K-ras siRNA transgene, providing additive tumor growth–inhibitory responses. Therapy with ONYX alone or ONXY-411 with green fluorescent protein (GFP) siRNA as controls had significantly lower therapeutic activity. The principles of antisense technology are conceptually simple. Oligonucleotides are designed to hybridize to a defined target mRNA and to inhibit its translation into protein.102,103 This approach was first used in 1978 by Stephenson and Zamecnik104 to inhibit the Rous sarcoma virus expression in chicken fibroblasts. Several antisense oligonucleotides are in clinical trials, and one has received FDA approval for the treatment of cytomegalovirus retinitis. Although it is relatively easy to synthesize phosphodiester oligonucleotides, they cannot be used as drugs because of their sensitivity to nuclease degradation. To improve their resistance to nuclease digestion, different chemical modifications are used, including phosphorothioates, methylphosphonates, and phosphoramidates.105 These modifications increase the stability of oligonucleotides, but they also alter the capacity to hybridize with RNA and reduce cellular internalization. Ribozymes are RNA molecules capable of sequence-specific cleavage of mRNA molecules.106 They selectively bind to target mRNAs, forming a duplex that is easily hydrolyzed, resulting in the suppression of specific genes.50 Two types of ribozymes, the hammerhead and hairpin, have been studied extensively.107 However, because the RNA backbone of ribozymes is an easy target for ribonucleases, they are biologically unstable in vivo50 and thus are used primarily for gene suppression and for the induction of apoptosis and inhibition of proliferation. Targeted gene therapy of cancer can be achieved through targeted gene expression and vector targeting.11,108,109 Although it is less important during ex vivo or IT gene delivery, targeted gene therapy becomes crucial with systemic gene transfer. Impediments to gene therapy include the poor selectivity of existing vectors and the low efficiency of gene transfer. Overcoming these hurdles are critical to achieving vectors that can be targeted and injected intravenously—an important goal given the systemic nature of cancer.
Gene Therapy in Oncology
Introduction
Vectors
Viral Gene Transfer Vectors
Retroviridae—Retrovirus
Recombinant Moloney Murine Leukemia Virus
Recombinant Lentivirus
Recombinant Adenovirus

Recombinant Adeno-Associated Virus
Recombinant Herpes Simplex Virus
Recombinant Pox Vectors
Recombinant Alphavirus Vectors (Sindbis)
Nonviral Gene Transfer Vectors
Direct DNA Injection/Transduction
Hydrodynamic Gene Delivery
Liposomes and Virosomes
Ballistic Delivery (Gene Gun)
Nanoparticles
Nucleic Acid–Based Therapeutics
DNA Transduction
RNA Transduction
Oligonucleotides
Small Interfering RNA
Antisense
Ribozymes
Gene Targeting
Gene Therapy in Oncology