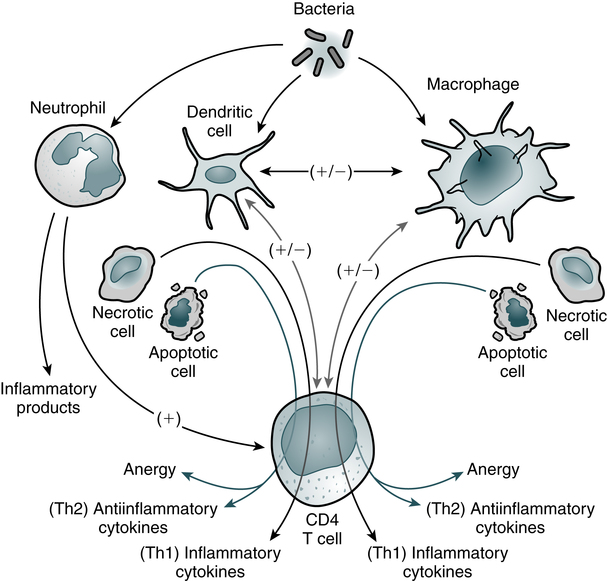
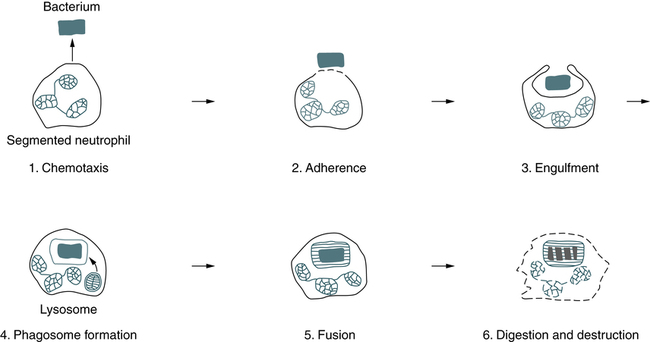
At the conclusion of this chapter, the reader should be able to: • Describe the general functions of granulocytes, monocytes-macrophages, and lymphocytes and plasma cells as components of the immune system. • Explain the process of phagocytosis. • Describe the composition and function of neutrophil extracellular traps (NETs). • Discuss the role of monocytes and macrophages in cellular immunity. • Define and compare acute inflammation and sepsis. • Briefly describe cell surface receptors. • Name and compare the signs and symptoms of disorders of neutrophil function. • Compare the signs and symptoms of two monocyte or macrophage disorders. • Describe states involving the leukocyte integrins. • Analyze case studies related to defects of neutrophils. • Correctly answer case study related multiple choice questions. • Be prepared to participate in a discussion of critical thinking questions. • Describe the principal reporting of results, sources of error, clinical applications, and limitations of a phagocytic engulfment test. The entire leukocytic cell system is designed to defend the body against disease. Each cell type has a unique function and behaves independently and, in many cases, in cooperation with other cell types. Leukocytes can be functionally divided into the general categories of granulocyte, monocyte-macrophage, and lymphocyte–plasma cell. The primary phagocytic cells are the polymorphonuclear neutrophil (PMN) leukocytes and the mononuclear monocytes-macrophages. The response of the body to pathogens involves cross-talk among many immune cells, including macrophages, dendritic cells, and CD4 T cells (Fig. 3-1). The lymphocytes participate in body defenses primarily through the recognition of foreign antigens and production of antibody. Plasma cells are antibody-synthesizing cells. 1. The first blood cells are primitive red blood cells (RBCs; erythroblasts) formed in the islets of the yolk sac during the first 2 to 8 weeks of life. 2. Gradually, the liver and spleen replace the yolk sac as the sites of blood cell development. By the second month of gestation, the liver becomes the major site of hematopoiesis, and granular types of leukocytes have made their initial appearance. The liver and spleen predominate from about 2 to 5 months of fetal life. 3. In the fourth month of gestation, bone marrow begins to produce blood cells. After the fifth fetal month, bone marrow begins to assume its ultimate role as the primary site of hematopoiesis. Neutrophilic leukocytes, particularly the polymorphonuclear (PMN) type (see Color Plate 4), provide an effective host defense against bacterial and fungal infections. The antimicrobial function of PMNs is essential in the innate immune response. Although the monocytes-macrophages and other granulocytes are also phagocytic cells, the PMN is the principal leukocyte associated with phagocytosis and a localized inflammatory response. The formation of an inflammatory exudate (pus), which develops rapidly in an inflammatory response, is composed primarily of neutrophils and monocytes. The granules of segmented neutrophils contain various antibacterial substances (Table 3-1). During the phagocytic process, the powerful antimicrobial enzymes that are released also disrupt the integrity of the cell itself. Neutrophils are also steadily lost to the respiratory, gastrointestinal (GI), and urinary systems, where they participate in generalized phagocytic activities. An alternate route for the removal of neutrophils from the circulation is phagocytosis by cells of the mononuclear phagocyte system. Table 3-1 Function and Types of Granules in Neutrophils Adapted from Peakman M, Vergani D: Basic and clinical immunology, ed 2, Edinburgh, 2009, Churchill Livingstone, p 24. The eosinophil (see Color Plate 5) is considered to be a homeostatic regulator of inflammation. Functionally, this means that the eosinophil attempts to suppress an inflammatory reaction to prevent the excessive spread of the inflammation. The eosinophil may also play a role in the host defense mechanism because of its ability to kill certain parasites. Basophils (see Color Plate 6) have high concentrations of heparin and histamine in their granules, which play an important role in acute, systemic, hypersensitivity reactions (see Chapter 26). Degranulation occurs when an antigen such as pollen binds to two adjacent immunoglobulin E (IgE) antibody molecules located on the surface of mast cells. The events resulting from the release of the contents of these basophilic granules include increased vascular permeability, smooth muscle spasm, and vasodilation. If severe, this reaction can result in anaphylactic shock. Phagocytosis can be divided into six stages—chemotaxis, adherence, engulfment, phagosome formation, fusion, and digestion and destruction (Fig. 3-2). The physical occurrence of damage to tissues, by trauma or microbial multiplication, releases substances such as activated complement components and products of infection to initiate phagocytosis. • Fc receptor—binds the Fc portion of antibody molecules, chiefly immunoglobulin G (IgG). The IgG attaches to the organism through its Fab site. • Complement receptor—the third component of complement, C3, also binds to organisms and then attaches to the complement receptor. 1. Antibody attached to the surface of a bacterium minimally binds the Fc phagocyte receptor. 2. Complement C3b is attached to the surface of the bacterium and binds loosely to the phagocyte C3b receptor. 3. Both antibody and C3b are attached to the surface of the bacterium and bound tightly to the phagocyte, allowing greater opportunity for the phagocyte to engulf the bacterium. On reaching the site of infection, phagocytes engulf and destroy the foreign matter (Fig. 3-3). Eosinophils can also undergo this process, except that they kill parasites. After the phagocytic cells have arrived at the site of injury, the bacteria can be engulfed through active membrane invagination. Pseudopodia are extended around the pathogen, pulled by interactions between the Fc receptors and Fc antibody portions on the opsonized bacterium. Pseudopodia meet and fuse, thereby internalizing the bacterium and enclosing it in a phagocytic vacuole, or phagosome.
Cells and Cellular Activities of the Immune System
Granulocytes and Mononuclear Cells
Origin and Development of Blood Cells
Granulocytic Cells
Neutrophils
Function
Azurophilic (Primary) Granules
Specific (Secondary) Granules
Microbicidal
Myeloperoxidase
Cytochrome b558 and other respiratory burst components
Lysozyme
Lysozyme
Elastase
Lactoferrin
Defensins
Cathepsin G
Proteinase-3
Bacterial permeability-increasing protein (BPI)
Cell migration
Collagenase
CD11b–CD18 (CR-3)
N-formulated peptides (e.g., N-formyl-methionyl-leucylphenylalanine receptor [FMLP-R])
Eosinophils and Basophils
Eosinophils
Basophils
Process of Phagocytosis
Chemotaxis
Engulfment
Cells and Cellular Activities of the Immune System: Granulocytes and Mononuclear Cells