Chapter 8 Antigen Presentation
• T cells survey proteins derived from intracellular or extracellular pathogens by recognizing peptide fragments that have been processed and become bound to major histocompatibility complex (MHC) class I or II molecules, respectively. These MHC–antigen complexes are presented at the cell surface.
• MHC class I molecules associate with endogenously synthesized peptides, produced by degradation of the cells’ internal molecules. This type of antigen processing is carried out by proteasomes, which cleave the proteins and transporters, which take the fragments to the endoplasmic reticulum (ER).
• MHC class II molecules bind to peptides produced following the breakdown of proteins that the cell has endocytosed. The peptides produced by degradation of these external antigens are loaded onto MHC class II molecules in a specialized endosomal compartment called MIIC.
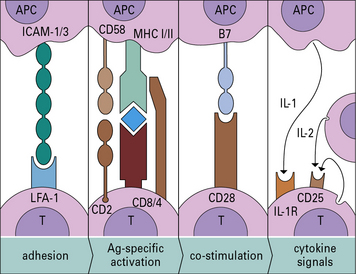
• The highly ordered area of contact between the T cell and APC is an immunological synapse. TCRs and costimulatory receptors occupy the center of the synapse. Adhesion molecules are found in the periphery.
• Costimulatory molecules are essential for T cell activation. Molecules such as B7 (CD80/86) on the APC bind to CD28 on the T cell to cause activation. Antigens presented without costimulation usually induce T cell anergy. Intercellular adhesion molecules also contribute to the interaction between a T cell and an antigen-presenting cell (APC). Interactions between intercellular cell adhesion molecule-1 (ICAM-1) and leukocyte functional antigen-1 (LFA-1) and between CD2 and its ligands extend the interaction between T cells and APCs.
• CD4 binds to MHC class II and CD8 to MHC class I molecules. These interactions increase the affinity of T cell binding to the appropriate MHC–antigen complex and bring kinases to the TCR complex.
• Binding of CTLA-4 or PD-1 on the T cell limits activation. Both of these ligands inhibit the costimulatory signal that the T cell receives from CD28.
• T cell activation induces enzyme cascades, leading to the production of interleukin-2 (IL-2) and the high-affinity IL-2 receptor on the T cell. IL-2 is required to drive T cell division.
• Antigen presentation affects the subsequent course of an immune response. The immune system responds to clues that an infection has taken place before responding strongly to antigens.
Antigen presenting cells
T cells only recognize antigen peptides bound to MHC-encoded molecules. Endogenous peptides, derived from intracellular sources such as replicating viruses, are presented on MHC class-I molecules to CD8+ T cells, while exogenous peptides, derived from extracellular sources such as microbes, are presented on MHC class-II molecules to CD4+ T cells. Before peptides can associate with the MHC molecules they are generated by partial proteolysis from the original protein antigen. Antigen processing refers to the degradation of antigen into peptide fragments, which may become bound to MHC class I or class II molecules (see Chapter 5). Whether a peptide binds to an MHC molecule depends on the amino-acid sequence of the peptide and on whether a suitable binding MHC molecule is available, which depends on the set of MHC molecules present in the individual. Broadly speaking, a single T cell recognizes a specific peptide(s) bound in the peptide-binding groove of a specific MHC molecule. However there are instances where T cells can respond to a different MHC/peptide combination, and this is equivalent to the cross-reactivity that may occur when antibodies bind to cross-reactive antigens.
• different types of antigen-presenting cell (APC) are brought into play depending on the situation. Dendritic cells (DCs) are particularly crucial for initiating responses;
• the interaction between the TCR, and the MHC/peptide complex is highly specific;
• another level of control is exerted by costimulatory molecules on APCs, resulting in T cell activation only when appropriate, such as in an infection;
• adhesion molecules on the interacting cells also contribute to the stable binding of the cells, which promotes effective antigen presentation;
• signals from the cell surface are then transmitted by a series of signal transduction pathways that regulate gene expression including cytokine production;
• in the final stages the actions of cytokines on the lymphocytes drive cell division.
The four stages of antigen presentation are outlined in Figure 8.1. In lymphoid organs, all four stages of the process can occur, resulting in T cell proliferation. However, antigen presentation can also occur to a more limited degree in tissues, resulting in cytokine production, but with little T cell division.
Interactions with antigen-presenting cells direct T cell activation
Dendritic cells are crucial for priming T cells
The majority of dendritic cells enter lymph nodes via afferent lymphatics. Originally it was thought that these cells were mostly derived from Langerhans’ cells in the skin, but it now appears that a substantial proportion of the early migrating DCs in afferent lymph are dermal dendritic cells (they do not express langerin (CD207) a marker of Langerhans’ cells). Moreover DCs derived from Langerhans’ cells tend to localize in the paracortex of the lymph node, whereas dermal DCs remain near lymphoid follicles. Dendritic cells arriving from the periphery of the body transport antigen to the lymph node and process it for presentation to T cells. A minor proportion of the DCs in lymph nodes, arrive from the blood across the HEV, using the same route as T cells and B cells (see Fig. 6.15), however these cells have not acquired antigen in the periphery and they can only acquire it from lymph or transfer from other cells. As they mature, dendritic cells express CCR7 which allows them to localize to the lymphoid tissues. There is also some evidence that DCs from skin and the gut have distinctive chemokine receptors, which allow them to selectively recirculate to their own lymphoid organs. As they mature, DCs also increase expression of key costimulatory molecules, including CD40, CD80 and CD86 (B7-1 and B7-2).
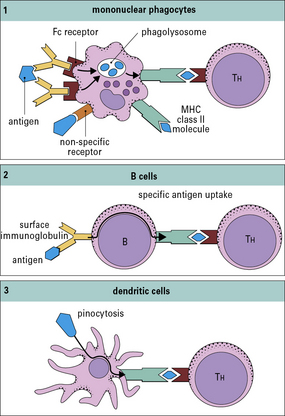
Macrophages and B cells present antigen to primed T cells
• bind to a specific antigen through surface IgM or IgD;
• then degrade it into peptides, which associate with MHC class II molecules.
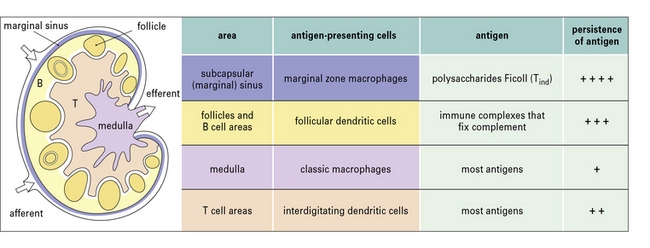
The properties and functions of some APCs are summarized in Figures 8.3 and 8.4.

Antigen processing
Antigen processing involves degrading the antigen into peptide fragments. The vast majority of epitopes recognized by T cells are fragments from a peptide chain. Only a minority of peptide fragments from a protein antigen are able to bind to a particular MHC molecule. Furthermore, different MHC molecules bind different sets of peptides (see Chapter 5). For example, the great majority of the immune response in humans against the HIV matrix protein is directed against a single immunodominant region, i.e. one which is recognized by a large number of T cells. However, exactly which part of this region is recognized, depends on the MHC haplotypes of the individual (Fig. 8.5).

MHC class I pathway
• influenza virus hemagglutinin (HA), a glycoprotein associated with the membrane of an infected host cell, normally elicits only a weak CTL response, but influenza virus HA can be generated in the cytoplasm by deleting that part of its sequence that encodes the N terminal signal peptide (required for translation across the membrane of the endoplasmic reticulum [ER]) and, when this is done, there is a strong CTL response to HA;
• the introduction of ovalbumin into the cytoplasm of a target cell (using an osmotic shock technique) generates CTLs recognizing ovalbumin, whereas the addition of exogenous ovalbumin generates an exclusively TH cell response.
Proteasomes are cytoplasmic organelles that degrade cytoplasmic proteins
Although the assembly of MHC class I molecules occurs in the ER of the cell, peptides destined to be presented by MHC class I molecules are generated from cytosolic proteins. The initial step in this process involves an organelle called the proteasome – a multi-protein complex which forms a barrel-like structure (Fig. 8.6).
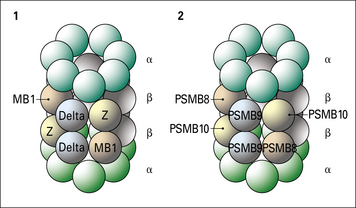
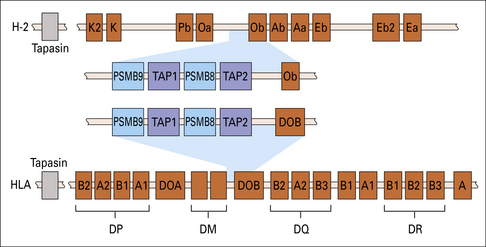
Two genes, PSMB8 and PSMB9 located in the class II region of the MHC (Fig. 8.7), encode proteasome components that subtly modify the range of peptides produced by proteasomes. The expression of these genes is induced by interferon-γ (IFNγ). The proteins displace constitutive subunits of the proteasome and along with a third inducible proteasome component (PSMB10 encoded on a different chromosome) influence processing of peptides by creating a wider range of peptide fragments suitable for binding MHC class I molecules. Additional subunits associate with the ends of core (20 S) proteasomes and may influence antigen processing. These include interferon-inducible PA28 (proteasome-activator-28) molecules as well as a complex of proteins that result in a larger 26 S particle.